Knotted protein
Knotted proteins are proteins whose backbones entangle themselves in a knot. One can imagine pulling a protein chain from both termini, as though pulling a string from both ends. When a knotted protein is “pulled” from both termini, it does not get disentangled. Knotted proteins are interesting because they are very rare, and their folding mechanisms and function are not well understood. Although there have been some experimental and theoretical studies that hinted to some answers, systematic answers to these questions have not been found.
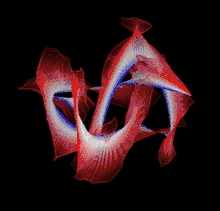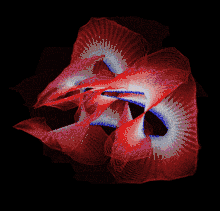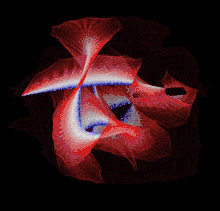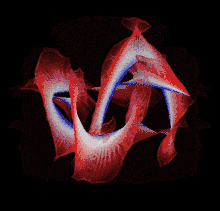
Though a number of computational methods have been developed for detecting protein knots, there are still no completely automatic methods to detect protein knots without the necessary manual intervention due to the missing residues or chain breaks in the X-ray structures or the nonstandard PDB formats.
Most of the knots discovered in proteins are deep trefoil (31) knots. Figure eight knots (41), three-twist knots (52), and Stevedore knots (61) have also been discovered.

Mathematical interpretation
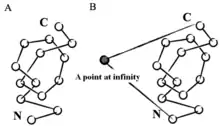
Mathematically, a knot is defined as a subset of three-dimensional space that is homeomorphic to a circle.[2] According to this definition, a knot only makes sense in a closed loop. However, many strategies have been used to create an artificial closed loop. For example, if we pick a point in space at infinite distance and connect it to the N and C termini through a virtual bond, the protein can be treated as a closed loop, or stochastic methods that create random closures.
Depth of the knot
The depth of a protein knot relates to the ability of the protein to resist unknotting. A deep knot is preserved even though the removal of a considerable number of residues from either end does not destroy the knot. The higher the number of residues that can be removed without destroying the knot, the deeper the knot.
Formation of knots
Considering how knots may be produced with a string, the folding of knotted proteins should involve first the formation of a loop, and then the threading of one terminus through the loop. This is the only topological way that the trefoil knot can be formed. For more complex knots, it is theoretically possible to have the loop to twist multiple times around itself, meaning that one end of the chain gets wrapped around at least once, and then threading to occur. It has also been observed in a theoretical study that a 6-1 knot can form by the C-terminus threading through a loop, and another loop flipping over the first loop, as well as the C-terminus threading through both the loops which have previously flipped over each other.[3]
There have been experimental studies involving YibK and YbeA, knotted proteins containing trefoil knots. It has been established that these knotted proteins fold slowly, and that the knotting in folding is the rate limiting step.[4] In another experimental study, a 91-residue-long protein was attached to the termini of YibK and YbeA.[5] Attaching the protein to both termini produces a deep knot with about 125 removable residues on each terminus before the knot is destroyed. Yet it was seen that the resulting proteins could fold spontaneously. The attached proteins were shown to fold more quickly than YibK and YbeA themselves, so during folding they are expected to act as plugs at either end of YibK and YbeA. It was found that attaching the protein to the N-terminus did not alter the folding speed, but the attachment to the C-terminus slows folding down, suggesting that the threading event happens at the C-terminus. The chaperones, although facilitate the protein knotting, are not crucial in proteins' self-tying.[5][6]
The folding of knotted proteins may be explained by interaction of the nascent chain with the ribosome. In particular, the affinity of the chain to the ribosome surface may result in creation of the loop, which may be next threaded by a nascent chain. Such mechanism was shown to be plausible for one of the most deeply knotted proteins known.[7]
Other topologically complex structures in proteins
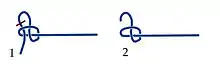
The class of knotted proteins contains only structures, for which the backbone, after closure forms a knotted loop. However, some proteins contain "internal knots" called slipknots, i.e. unknotted structures containing a knotted subchain.[8] Another topologically complex structure is the link formed by covalent loops, closed by disulfide bridges.[9][10] Three types of disulfide-based links were identified in proteins: two versions of Hopf link (differing in chirality) and one version of Solomon link. Another complex structure arising by closing part of the chain with covalent bridge are complex lasso proteins, for which the covalent loop is threaded by the chain one or more times.[11] Yet another complex structures arising as a result of the existence of disulfide bridges are the cystine knots, for which two disulfide bridges form a closed, covalent loop, which is threaded by third chain. The term "knot" in the name of the motif is misleading, as the motif does not contain any knotted closed cycle. Moreover, formation of the cystine knots in general is not different from the folding of an unknotted protein
Apart from closing only one chain, one may perform also the chain closure procedure for all the chains present in the crystal structure. In some cases one obtains the non-trivially linked structures, called probabilistic links.[12]
One can also consider loops in proteins formed by pieces of the main chain and the disulfide bridges and interaction via ions. Such loops can also be knotted of form links even within structures with unknotted main chain.[13][14]
First discoveries
Marc L. Mansfield proposed in 1994, that there can be knots in proteins.[15] He gave unknot scores to proteins by constructing a sphere centered at the center of mass of the alpha carbons of the backbone, with a radius twice the distance between the center of mass and the Calpha that is the farthest away from the center of mass, and by sampling two random points on the surface of the sphere. He connected the two points by tracing a geodesic on the surface of the sphere (arcs of great circles), and then connected each end of the protein chain with one of these points. Repeating this procedure a 100 times and counting the times where the knot is destroyed in the mathematical sense yields the unknot score. Human carbonic anhydrase was identified to have a low unknot score (22). Upon visually inspecting the structure, it was seen that the knot was shallow, meaning that the removal of a few residues from either end destroys the knot.
In 2000, William R. Taylor identified a deep knot in acetohydroxy acid isomeroreductase (PDB ID: 1YVE), by using an algorithm that smooths protein chains and makes knots more visible.[16] The algorithm keeps both termini fixed, and iteratively assigns to the coordinates of each residue the average of the coordinates of the neighboring residues. It has to be made sure that the chains do not pass through each other, otherwise the crossings and therefore the knot might get destroyed. If there is no knot, the algorithm eventually produces a straight line that joins both termini.
Studies about the function of the knot in a protein
Some proposals about the function of knots have been that it might increase thermal and kinetic stability. One particular suggestion was that for the human ubiquitin hydrolase, which contains a 5-2 knot, the presence of the knot might be preventing it from being pulled into the proteasome.[17] Because it is a deubiquitinating enzyme, it is often found in proximity of proteins soon to be degraded by proteasome, and therefore it faces the danger of being degraded itself. Therefore, the presence of the knot might be functioning as a plug that prevents it. This notion was further analyzed on other proteins like YbeA and YibK with computer simulations.[18] The knots seem to tighten when they are pulled into a pore, and depending on the force with which they are pulled in, they either get stuck and block the pore, the likeliness of which increases with stronger pulling forces, or in the case of a small pulling force they might get disentangled as one terminus is pulled out of the knot. For deeper knots, it is more likely that the pore will be blocked, as there are too many residues that need to be pulled through the knot. In another theoretical study,[19] it was found that the modeled knotted protein was not thermally stable, but it was kinetically stable. It was also shown, that the knot in proteins creates the places on the verge of hydrophobic and hydrophilic parts of the chain, characteristic for active sites.[20] This may explain, why over 80% of knotted proteins are enzymes.[21]
Web servers to extrapolate knotted proteins
Some local programs[22] and a number of web servers are available, providing convenient query services for knotted structures and analysis tools for detecting protein knots.[14][21][23]
See also
References
- Robert, Scharein. "KnotPlot: Hypnagogic Software (Version 0.1)". Nearly all of the images here were created with KnotPlot, a fairly elaborate program to visualize and manipulate mathematical knots in three and four dimensions.
- Cromwell, P. D. (2004). Knots and Links. Cambridge: Cambridge University Press.
- Bölinger, D.; Sułkowska, J.I.; Hsu, H-P.; Mirny, L.A.; Kardar, M. (1 April 2010). "A Stevedore's Protein Knot". PLOS Comput Biol. 6 (4): e1000731. Bibcode:2010PLSCB...6E0731B. doi:10.1371/journal.pcbi.1000731. PMC 2848546. PMID 20369018.
- Mallam, A.L.; Jackson, S.E. (2012). "Knot formation in newly translated proteins is spontaneous and accelerated by chaperonins". Nat Chem Biol. 8 (2): 147–153. doi:10.1038/nchembio.742. PMID 22179065.
- Lim, Nicole C.H.; Jackson, S.E. (30 January 2015). "Mechanistic insights into the folding of knotted proteins in vitro and in vivo". J. Mol. Biol. 427 (2): 248–258. doi:10.1016/j.jmb.2014.09.007. PMID 25234087.
- Zhao, Yani; Dabrowski-Tumanski, Pawel; Niewieczerzal, Szymon; Sulkowska, Joanna I. (2018-03-16). "The exclusive effects of chaperonin on the behavior of proteins with 52 knot". PLOS Computational Biology. 14 (3): e1005970. Bibcode:2018PLSCB..14E5970Z. doi:10.1371/journal.pcbi.1005970. ISSN 1553-7358. PMC 5874080. PMID 29547629.
- Dabrowski-Tumanski, Pawel; Piejko, Maciej; Niewieczerzal, Szymon; Stasiak, Andrzej; Sulkowska, Joanna I. (2018-10-12). "Protein Knotting by Active Threading of Nascent Polypeptide Chain Exiting from the Ribosome Exit Channel". The Journal of Physical Chemistry B. 122 (49): 11616–11625. doi:10.1021/acs.jpcb.8b07634. ISSN 1520-6106. PMID 30198720.
- King, Neil P.; Yeates, Eric O.; Yeates, Todd O. (October 2007). "Identification of Rare Slipknots in Proteins and Their Implications for Stability and Folding". Journal of Molecular Biology. 373 (1): 153–166. doi:10.1016/j.jmb.2007.07.042. ISSN 0022-2836. PMID 17764691.
- Dabrowski-Tumanski, Pawel; Sulkowska, Joanna I. (2017-03-28). "Topological knots and links in proteins". Proceedings of the National Academy of Sciences. 114 (13): 3415–3420. doi:10.1073/pnas.1615862114. ISSN 0027-8424. PMC 5380043. PMID 28280100.
- Boutz, Daniel R.; Cascio, Duilio; Whitelegge, Julian; Perry, L. Jeanne; Yeates, Todd O. (May 2007). "Discovery of a Thermophilic Protein Complex Stabilized by Topologically Interlinked Chains". Journal of Molecular Biology. 368 (5): 1332–1344. doi:10.1016/j.jmb.2007.02.078. ISSN 0022-2836. PMC 1955483. PMID 17395198.
- Niemyska, Wanda; Dabrowski-Tumanski, Pawel; Kadlof, Michal; Haglund, Ellinor; Sułkowski, Piotr; Sulkowska, Joanna I. (2016-11-22). "Complex lasso: new entangled motifs in proteins". Scientific Reports. 6 (1): 36895. Bibcode:2016NatSR...636895N. doi:10.1038/srep36895. ISSN 2045-2322. PMC 5118788. PMID 27874096.
- Dabrowski-Tumanski, Pawel; Jarmolinska, Aleksandra I.; Niemyska, Wanda; Rawdon, Eric J.; Millett, Kenneth C.; Sulkowska, Joanna I. (2016-10-28). "LinkProt: a database collecting information about biological links". Nucleic Acids Research. 45 (D1): D243–D249. doi:10.1093/nar/gkw976. ISSN 0305-1048. PMC 5210653. PMID 27794552.
- Liang, Chengzhi; Mislow, Kurt (November 1994). "Knots in Proteins". Journal of the American Chemical Society. 116 (24): 11189–11190. doi:10.1021/ja00103a057. ISSN 0002-7863.
- Dabrowski-Tumanski, Pawel; Rubach, Pawel; Goundaroulis, Dimos; Dorier, Julien; Sułkowski, Piotr; Millett, Kenneth C.; Rawdon, Eric J.; Stasiak, Andrzej; Sulkowska, Joanna I. (2018). "KnotProt 2.0: a database of proteins with knots and other entangled structures". Nucleic Acids Research. 47 (D1): D367–D375. doi:10.1093/nar/gky1140. PMC 6323932. PMID 30508159.
- Mansfield, Marc L. (1994). "Are there knots in proteins?". Nat. Struct. Biol. 1 (4): 213–214. doi:10.1038/nsb0494-213. PMID 7656045. S2CID 32625699.
- Taylor, William R. (2000). "A deeply knotted protein structure and how it might fold". Nature. 406 (6798): 916–919. Bibcode:2000Natur.406..916T. doi:10.1038/35022623. PMID 10972297. S2CID 4420225.
- Virnau, Peter; Mirny, L.A.; Kardar, M. (2006). "Intricate knots in proteins: function and evolution". PLOS Comput Biol. 2 (9): e122. Bibcode:2006PLSCB...2..122V. doi:10.1371/journal.pcbi.0020122. PMC 1570178. PMID 16978047.
- Szymczak, P. (2014). "Translocation of knotted proteins through a pore". European Physical Journal ST. 223 (9): 1805–1812. Bibcode:2014EPJST.223.1805S. doi:10.1140/epjst/e2014-02227-6. S2CID 16379224.
- Soler, M.A.; Nunes, A.; Faisca, P. F. N. (2014). "Effects of knot type in the folding of topologically complex lattice proteins". J. Chem. Phys. 141 (2): 025101. Bibcode:2014JChPh.141b5101S. doi:10.1063/1.4886401. PMID 25028045.
- Dabrowski-Tumanski, Pawel; Stasiak, Andrzej; Sulkowska, Joanna I. (2016-11-02). "In Search of Functional Advantages of Knots in Proteins". PLOS ONE. 11 (11): e0165986. Bibcode:2016PLoSO..1165986D. doi:10.1371/journal.pone.0165986. ISSN 1932-6203. PMC 5091781. PMID 27806097.
- Jamroz, M; Niemyska W; Rawdon EJ; Stasiak A; Millett KC; Sułkowski P; Sulkowska JI (2015). "KnotProt: a database of proteins with knots and slipknots". Nucleic Acids Research. 43 (Database): D306–D314. doi:10.1093/nar/gku1059. PMC 4383900. PMID 25361973.
- Jarmolinska, AI; Gambin A; Sulkowska JI (2019). "Knot_pull—python package for biopolymer smoothing and knot detection". Bioinformatics. 36 (3): 953–955. doi:10.1093/bioinformatics/btz644. PMID 31504154.
- Lai, Y.-L.; Yen, S.-C.; Yu, S.-H.; Hwang, J.-K. (7 May 2007). "pKNOT: the protein KNOT web server". Nucleic Acids Research. 35 (Web Server): W420–W424. doi:10.1093/nar/gkm304. PMC 1933195. PMID 17526524.