Non-photochemical quenching
Non-photochemical quenching (NPQ) is a mechanism employed by plants and algae to protect themselves from the adverse effects of high light intensity.[1] It involves the quenching of singlet excited state chlorophylls (Chl) via enhanced internal conversion to the ground state (non-radiative decay), thus harmlessly dissipating excess excitation energy as heat through molecular vibrations. NPQ occurs in almost all photosynthetic eukaryotes (algae and plants), and helps to regulate and protect photosynthesis in environments where light energy absorption exceeds the capacity for light utilization in photosynthesis.[2]
Process
When a molecule of chlorophyll absorbs light it is promoted from its ground state to its first singlet excited state. The excited state then has three main fates. Either the energy is; 1. passed to another chlorophyll molecule by Förster resonance energy transfer (in this way excitation is gradually passed to the photochemical reaction centers (photosystem I and photosystem II) where energy is used in photosynthesis (called photochemical quenching)); or 2. the excited state can return to the ground state by emitting the energy as heat (called non-photochemical quenching); or 3. the excited state can return to the ground state by emitting a photon (fluorescence).
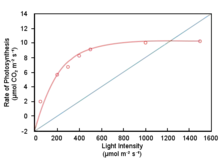
In higher plants, the absorption of light continues to increase as light intensity increases, while the capacity for photosynthesis tends to saturate. Therefore, there is the potential for the absorption of excess light energy by photosynthetic light harvesting systems. This excess excitation energy leads to an increase in the lifetime of singlet excited chlorophyll, increasing the chances of the formation of long-lived chlorophyll triplet states by inter-system crossing. Triplet chlorophyll is a potent photosensitiser of molecular oxygen forming singlet oxygen which can cause oxidative damage to the pigments, lipids and proteins of the photosynthetic thylakoid membrane. To counter this problem, one photoprotective mechanism is so-called non-photochemical quenching (NPQ), which relies upon the conversion and dissipation of the excess excitation energy into heat. NPQ involves conformational changes within the light harvesting proteins of photosystem (PS) II that bring about a change in pigment interactions causing the formation of energy traps. The conformational changes are stimulated by a combination of transmembrane proton gradient, the photosystem II subunit S (PsBs) and the enzymatic conversion of the carotenoid violaxanthin to zeaxanthin (the xanthophyll cycle).
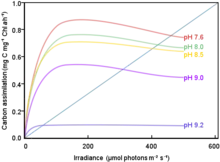
Violaxanthin is a carotenoid downstream from chlorophyll a and b within the antenna of PS II and nearest to the special chlorophyll a located in the reaction center of the antenna. As light intensity increases, acidification of the thylakoid lumen takes place through the stimulation of carbonic anhydrase, which in turn converts bicarbonate (HCO3) into carbon dioxide causing an influx of C02 and inhibiting Rubisco oxygenase activity.[5] This acidification also leads to the protonation of the PsBs subunit of PS II which catalyze the conversion of violaxanthin to zeaxanthin, and is involved in the alteration orientation of the photosystems at times of high light absorption to reduce the quantities of carbon dioxide created and start the non-photochemical quenching, along with the activation of enzyme violaxanthin de-epoxidase which eliminates an epoxide and forms an alkene on a six-member ring of violaxanthin giving rise to another carotenoid known as antheraxanthin. Violaxanthin contains two epoxides each bonded to a six-member ring and when both are eliminated by de-epoxidase the carotenoid zeaxanthin is formed. Only violaxanthin is able to transport a photon to the special chlorophyll a. Antheraxanthin and zeaxanthin dissipate the energy from the photon as heat preserving the integrity of photosystem II. This dissipation of energy as heat is one form of non-photochemical quenching.[6]
Measurement of NPQ
Non-photochemical quenching is measured by the quenching of chlorophyll fluorescence and is distinguished from photochemical quenching by applying a bright light pulse to transiently saturate photochemical quenching thus removing its contribution from the observed quenching. Non-photochemical quenching is not affected if the pulse of light is short. During this pulse, the fluorescence reaches the level reached in the absence of any photochemical quenching, known as maximum fluorescence, .

For further discussion, see Measuring chlorophyll fluorescence and Plant stress measurement.
Chlorophyll fluorescence can easily be measured with a chlorophyll fluorometer. Some fluorometers can calculate NPQ and photochemical quenching coefficients (including qP, qN, qE and NPQ), as well as light and dark adaptation parameters (including Fo, Fm, and Fv/Fm).
References
- Horton, Peter; Alexander V. Ruban (April 2005). "Regulation of Photosynthesis under Stress: Molecular design of the photosystem II light-harvesting antenna: photosynthesis and photoprotection". J. Exp. Bot. 56 (411): 365–373. doi:10.1093/jxb/eri023. PMID 15557295.
- Müller, Patricia; Xiao-Ping Li; Krishna K. Niyogi (April 2001). "Update on Photosynthesis: Non-Photochemical Quenching. A Response to Excess Light Energy". Plant Physiol. 125 (4): 1558–1566. doi:10.1104/pp.125.4.1558. PMC 1539381. PMID 11299337.
- Masahiro Tamoi; Miki Nagaoka; Yoshiko Miyagawa; Shigeru Shigeoka (2006). "Contribution of Fructose-1,6-bisphosphatase and Sedoheptulose-1,7-bisphosphatase to the Photosynthetic Rate and Carbon Flow in the Calvin Cycle in Transgenic Plants". Plant & Cell Physiology. 29 (10): 380–390. doi:10.1093/pcp/pcj004. PMID 16415064.
- Kristian Spilling (2007). "Dense sub-ice bloom of dinoflagellates in the Baltic Sea, potentially limited by high pH". Journal of Plankton Research. 29 (10): 895–901. doi:10.1093/plankt/fbm067.
- Raven, John Albert (June 2008). "CO2-concentrating mechanisms: A direct role for thylakoid lumen acidification". Plant, Cell & Environment. 20 (2): 147–154. doi:10.1046/j.1365-3040.1997.d01-67.x. Retrieved November 20, 2020.
- Baker, Neil R. (2008-01-01). "Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo". Annual Review of Plant Biology. 59 (1): 89–113. doi:10.1146/annurev.arplant.59.032607.092759. PMID 18444897.