Photoactivatable probes
Photoactivation is a technique used in biological research to specifically activate cellular players (proteins, nucleic acids, small molecules) by a flash of light in order to study processes in cells. The basic principle is to bring a photoactivatable agent (e.g. a small molecule modified with a light-responsive group, proteins tagged with an artificial photoreceptor protein) to cells, tissue or even living animals and specifically control the activity by illumination [1]
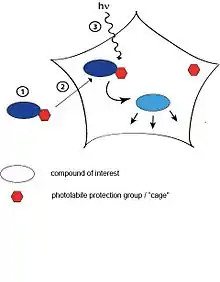
Light is a well-suited external trigger for these types of experiments since it is non-invasive and does not influence normal cellular processes (care has to be taken when using light in the ultra-violet part of the spectrum to avoid DNA damage. Furthermore, light offers high spatial and temporal control. Usually, the activation stimulus comes from a laser or a UV lamp and can be incorporated into the same microscope used for monitoring of the effect. All these advantages have led to the development of a wide variety of different photoactivatable probes.
Even though the light-induced activation step is usually irreversible, reversible changes can be induced in a number of photoswitches, which will not be discussed in detail here.
History
The first reported use of photoprotected analogues for biological studies was the synthesis and application of ‘caged’ ATP by Hoffman in 1978[2] in his study of Na:K pumps. To this day, ATP is still the most commonly used caged compound. Hoffman was also the one to coin the term 'caged compound' for this type of modified molecules. This nomenclature continued to persist, even if it is not scientifically correct. It suggests the image of a molecule in a physical cage (like in a Fullerene), therefore scientists have tried to introduce the newer, more accurate term "photoactivatable probes". Both nomenclatures are currently in use. Small molecules are easier to modify by photocleavable groups, compared to larger constructs such as proteins. Major discoveries were made in the following years with caged neurotransmitters such as glutamate, which is used to map functional neuronal circuits in mammalian brain slices.[3] Photoactivatable proteins were serendipitously discovered much later (in 2002), by the observation that Kaede protein, when left on the bench exposed to sunlight, changed fluorescence to longer wavelength. (for an extensive review, visit:[4])
Photoactivatable proteins
Proteins which sense and react to light were originally isolated from photoreceptors in algae, corals and other marine organisms. The two most commonly used photoactivatable proteins in science today are photoactivatable fluorescent proteins and retinylidene proteins. Photoactivatable fluorescent proteins change to longer emission wavelength upon illumination with UV light. In Kaede, this change is brought upon by cleavage of the chromophore tripeptide His62-Tyr63-Gly64.[5] This discovery paved the way for modern super resolution microscopy techniques like PALM or STORM. Retinylidene proteins such as Channelrhodopsins or Halorhodopsins are light sensitive cation and chloride channels, which open during illumination with blue and yellow light, respectively. This principle has been successfully employed to control the activity of neurons in living cells and even tissue and gave rise to a whole new research field, optogenetics.
Photoactivatable nucleic acids
Nucleic acids play important roles as cellular information storage and gene regulation machinery. In efforts to regulate this machinery by light, DNA and RNA have been modified with photocleavable groups at the backbone (in an approach called ‘statistical backbone caging’, the protection groups react mainly with backbone phosphate groups). In the organism, modified nucleic acids are ‘silent’ and only upon irradiation with light can their activity be turned on.[6] This approach finds use in developmental biology, where the chronology of gene activity is of particular interest. It is now possible to very precisely turn on genes of interest during the development of whole organisms.[7]
Photoactivatable small molecules
Small molecules are easily modified by chemical synthesis and therefore were among the first to be modified and used in biological studies. Until today, a wide variety of caged small molecules exist such that only a small representative section will be discussed here. A field where all the advantages of activating effectors with light (precise control, fast response, high specificity, no cross-reactions) are particularly interesting in the study of neurotransmitters.
Caged neurotransmitters
Caged dopamine, serotonin, glycine and GABA have been synthesized and their effect on neuronal activity has been extensively studied.[8]
Caged ions
Not only amino acids, but also ions can be caged. Since calcium is a potent cellular second messenger, caged variants have been synthesized by employing the ion-trapping properties of EDTA. Light-induced cleavage of the EDTA backbone leads to a wave of free calcium inside the cell.[9]
Caged hormones
Another class of molecules used for transmitting signals in the cell is hormones. Caged derivates of estradiol were shown to induce gene expression upon uncaging other caged hormones were used to study receptor – ligand interactions.[10]
Caged lipids
Even though lipids were long thought to just be building blocks of cellular membranes, it is now becoming clear, that some of them have a specific function also in signaling To dissect the roles that lipids have in certain pathways, it is advantageous to be able to increase the concentration of the signaling lipid in a very rapid manner. Therefore, many signaling lipids have been also protected with photoremovable protection groups and their effect on cellular signaling has been studied. Caged PI3P has been shown to induce endosomal fusion.[11] Caged IP3 helped elucidate the effect of IP3 on the action potential of neurons[12] and caged diacylglycerol has been used to determine the influence of fatty acid chain length on PKC dependent signaling.[13]
Photoactivatable lipids
When studying protein-lipid interactions, another type of photoactivation has proved to provide many insights. Photolabile groups such as diaziridines or benzophenones, which, upon UV irradiation leave behind a highly reactive carbenium ions, can be used to crosslink the lipid of interest to its interacting proteins. This methodology is especially useful to verify existing and discover new protein-lipid interactions.[14]
See also
References
- Rana, A.; Dolmetsch, R. E. (2010). "Using light to control signaling cascades in live neurons". Curr Opin Neurobiol. 20 (5): 617–622. doi:10.1016/j.conb.2010.08.018. PMC 2993759. PMID 20850295.
- Kaplan, J. H.; Forbush, B.; Hoffman, J. F. (1978). "Rapid photolytic release of adenosine 5'-triphosphate from a protected analogue: utilization by the Na:K pump of human red blood cell ghosts". Biochemistry. 17 (10): 1929–35. doi:10.1021/bi00603a020.
- Callaway, E. M.; Katz, L. C. (1993). "Photoactivation with caged glutamate reveals functional circuitry in living brain slices". Proceedings of the National Academy of Sciences of the United States of America. 90 (16): 7661–5. Bibcode:1993PNAS...90.7661C. doi:10.1073/pnas.90.16.7661. PMC 47202. PMID 7689225.
- Chudakov, D.; Matz, M. (2010). "Fluorescent proteins and their applications in imaging living cells and tissues". Physiological Reviews. 90 (3): 1103–1163. doi:10.1152/physrev.00038.2009. PMID 20664080.
- Tsutsui, H; Shimizu, H; Mizuno, H; Nukina, N; Furuta, T; Miyawaki, A (Nov 2009). "The E1 mechanism in photo-induced beta-elimination reactions for green-to-red conversion of fluorescent proteins". Chem Biol. 16: 1140–7. doi:10.1016/j.chembiol.2009.10.010. PMID 19942137.
- Mayer, G.; Heckel, A. (2006). "Biologically active molecules with a "light switch"". Angewandte Chemie International Edition in English. 45 (30): 4900–21. doi:10.1002/anie.200600387. PMID 16826610.
- Ando, H.; Futura, T.; Okamoto, H. (2004). "Photo-mediated gene activation by using caged mRNA in zebrafish embryos". Methods Cell Biol. 77: 159–71.
- Kramer, R.H.; Fortin, D.L.; Trauner, D. (2009). "New photochemical tools for controlling neuronal activity". Curr Opin Neurobiol. 19 (5): 544–52. doi:10.1016/j.conb.2009.09.004. PMC 2788492. PMID 19828309.
- Ellies-Davies, G.C. (2008). "Neurobiology with caged calcium". Chem. Rev. 108 (5): 1603–13. doi:10.1021/cr078210i. PMID 18447376.
- Link, K.H.; Cruz, F.G.; Ye, H.F.; O'reilly, K.E.; Dowdell, S.; Koh, J.T. (2004). "Photo-caged agonists of the nuclear receptors RARgamma and TRbeta provide unique time-dependent gene expression profiles for light-activated gene patterning". Bioorg. Med. Chem. 12 (22): 5949–59. doi:10.1016/j.conb.2009.09.004. PMC 2788492. PMID 19828309.
- Subramanian, D.; Laketa, V.; Müller, R.; Tischer, C.; Zarbakhsh, S.; Pepperkok, R.; Schultz, C. (2010). "Activation of membrane-permeant caged PtdIns(3)P induces endocomal fusion in cells". Nat Chem Biol. 6 (5): 324–6. doi:10.1038/nchembio.348.
- Lemtiri-Chlieh, F.; MacRobbie, E.A.; Brearley, C.A. (2000). "Inositol hexakisphosphate is a physiological signal regulating the K+-inward rectifying conductance in guard cells". Proc Natl Acad Sci USA. 97 (15): 8687–92. Bibcode:2000PNAS...97.8687L. doi:10.1073/pnas.140217497. PMC 27009. PMID 10890897.
- Nadler, A.; Reither, G.; Feng, S.; Stein, F.; Reither, S.; Müller, R.; Schultz, C. (2013). "The fatty acid composition of diacylglyerols determines local signaling patterns". Angew Chem Int Ed. 52: 6330–6334. doi:10.1002/anie.201301716.
- Haberkant, P.; Raijmakers, R.; Wildwater, M.; Sachsenheimer, T.; Brügger, B.; Maeda, K.; Houweling, M.; Gavin, A.C.; Schultz, C.; van Meer, G.; Heck, A.J.; Holthuis, J.C. (2013). "In vivo profiling and visualization of cellular protein-lipid interactions using bifunctional fatty acids". Angew Chem Int Ed Engl. 52 (14): 4033–8. doi:10.1002/anie.201210178.
External links
| Library resources about Photoactivatable probes |