Progesterone
Progesterone (P4) is an endogenous steroid and progestogen sex hormone involved in the menstrual cycle, pregnancy, and embryogenesis of humans and other species.[1][13] It belongs to a group of steroid hormones called the progestogens,[13] and is the major progestogen in the body. Progesterone has a variety of important functions in the body. It is also a crucial metabolic intermediate in the production of other endogenous steroids, including the sex hormones and the corticosteroids, and plays an important role in brain function as a neurosteroid.[14]
 | |
 | |
| Names | |
|---|---|
| IUPAC name
(8S,9S,10R,13S,14S,17S)-17-acetyl-10,13-dimethyl-1,2,6,7,8,9,11,12,14,15,16,17-dodecahydrocyclopenta[a]phenanthren-3-one | |
| Other names | |
| Identifiers | |
3D model (JSmol) |
|
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| DrugBank | |
| ECHA InfoCard | 100.000.318 |
| KEGG | |
PubChem CID |
|
| UNII | |
CompTox Dashboard (EPA) |
|
| |
| |
| Properties | |
| C21H30O2 | |
| Molar mass | 314.469 g/mol |
| Melting point | 126 |
| log P | 4.04[4] |
| Pharmacology | |
| G03DA04 (WHO) | |
| By mouth, topical/transdermal, vaginal, intramuscular injection, subcutaneous injection, subcutaneous implant | |
| Pharmacokinetics: | |
| OMP: <10%[5][6] | |
| • Albumin: 80% • CBG: 18% • SHBG: <1% • Free: 1–2%[7][8] | |
| Hepatic (CYP2C19, CYP3A4, CYP2C9, 5α-reductase, 3α-HSD, 17α-hydroxylase, 21-hydroxylase, 20α-HSD)[9][10] | |
| OMP: 16–18 hours[5][6][11] IM: 22–26 hours[6][12] SC: 13–18 hours[12] | |
| Renal | |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
| Infobox references | |
In addition to its role as a natural hormone, progesterone is also used as a medication, such as in combination with estrogen for contraception, to reduce the risk of uterine or cervical cancer, and in menopausal hormone therapy and hormone replacement therapy (HRT) in cases where a woman’s ovaries have been removed due to cancer or other physical issue.[15] It was first prescribed in 1934.[16]
Biological activity
Progesterone is the most important progestogen in the body. As a potent agonist of the nuclear progesterone receptor (nPR) (with an affinity of KD = 1 nM) the resulting effects on ribosomal transcription plays a major role in regulation of female reproduction.[13][17][18] In addition, progesterone is an agonist of the more recently discovered membrane progesterone receptors (mPRs),[19] of which the expression has regulation effects in reproduction function (oocyte maturation, labor, and sperm motility) and cancer although additional research is required to further define the roles.[20] It also functions as a ligand of the PGRMC1 (progesterone receptor membrane component 1) which impacts tumor progression, metabolic regulation, and viability control of nerve cells.[21][22][23] Moreover, progesterone is also known to be an antagonist of the sigma σ1 receptor,[24][25] a negative allosteric modulator of nicotinic acetylcholine receptors,[14] and a potent antagonist of the mineralocorticoid receptor (MR).[26] Progesterone prevents MR activation by binding to this receptor with an affinity exceeding even those of aldosterone and glucocorticoids such as cortisol and corticosterone,[26] and produces antimineralocorticoid effects, such as natriuresis, at physiological concentrations.[27] In addition, progesterone binds to and behaves as a partial agonist of the glucocorticoid receptor (GR), albeit with very low potency (EC50 >100-fold less relative to cortisol).[28][29]
Progesterone, through its neurosteroid active metabolites such as 5α-dihydroprogesterone and allopregnanolone, acts indirectly as a positive allosteric modulator of the GABAA receptor.[30]
Progesterone and some of its metabolites, such as 5β-dihydroprogesterone, are agonists of the pregnane X receptor (PXR),[31] albeit weakly so (EC50 >10 μM).[32] In accordance, progesterone induces several hepatic cytochrome P450 enzymes,[33] such as CYP3A4,[34][35] especially during pregnancy when concentrations are much higher than usual.[36] Perimenopausal women have been found to have greater CYP3A4 activity relative to men and postmenopausal women, and it has been inferred that this may be due to the higher progesterone levels present in perimenopausal women.[34]
Progesterone modulates the activity of CatSper (cation channels of sperm) voltage-gated Ca2+ channels. Since eggs release progesterone, sperm may use progesterone as a homing signal to swim toward eggs (chemotaxis). As a result, it has been suggested that substances that block the progesterone binding site on CatSper channels could potentially be used in male contraception.[37][38]
Biological function
Hormonal interactions
Progesterone has a number of physiological effects that are amplified in the presence of estrogens. Estrogens through estrogen receptors (ERs) induce or upregulate the expression of the PR.[39] One example of this is in breast tissue, where estrogens allow progesterone to mediate lobuloalveolar development.[40][41][42]
Elevated levels of progesterone potently reduce the sodium-retaining activity of aldosterone, resulting in natriuresis and a reduction in extracellular fluid volume. Progesterone withdrawal, on the other hand, is associated with a temporary increase in sodium retention (reduced natriuresis, with an increase in extracellular fluid volume) due to the compensatory increase in aldosterone production, which combats the blockade of the mineralocorticoid receptor by the previously elevated level of progesterone.[43]
Reproductive system

Progesterone has key effects via non-genomic signalling on human sperm as they migrate through the female tract before fertilization occurs, though the receptor(s) as yet remain unidentified.[44] Detailed characterisation of the events occurring in sperm in response to progesterone has elucidated certain events including intracellular calcium transients and maintained changes,[45] slow calcium oscillations,[46] now thought to possibly regulate motility.[47] It is produced by the ovaries.[48] Progesterone has also been shown to demonstrate effects on octopus spermatozoa.[49]
Progesterone is sometimes called the "hormone of pregnancy",[50] and it has many roles relating to the development of the fetus:
- Progesterone converts the endometrium to its secretory stage to prepare the uterus for implantation. At the same time progesterone affects the vaginal epithelium and cervical mucus, making it thick and impenetrable to sperm. Progesterone is anti-mitogenic in endometrial epithelial cells, and as such, mitigates the tropic effects of estrogen.[51] If pregnancy does not occur, progesterone levels will decrease, leading, in the human, to menstruation. Normal menstrual bleeding is progesterone-withdrawal bleeding. If ovulation does not occur and the corpus luteum does not develop, levels of progesterone may be low, leading to anovulatory dysfunctional uterine bleeding.
- During implantation and gestation, progesterone appears to decrease the maternal immune response to allow for the acceptance of the pregnancy.[52]
- Progesterone decreases contractility of the uterine smooth muscle.[50] This effect contributes to prevention of preterm labor.[52]
- A drop in progesterone levels is possibly one step that facilitates the onset of labor.
- In addition progesterone inhibits lactation during pregnancy. The fall in progesterone levels following delivery is one of the triggers for milk production.
The fetus metabolizes placental progesterone in the production of adrenal steroids.
Breasts
Lobuloalveolar development
Progesterone plays an important role in breast development in women. In conjunction with prolactin, it mediates lobuloalveolar maturation of the mammary glands during pregnancy to allow for milk production and thus lactation and breastfeeding of offspring following parturition (childbirth).[53] Estrogen induces expression of the PR in breast tissue and hence progesterone is dependent on estrogen to mediate lobuloalveolar development.[40][41][42] It has been found that RANKL is a critical downstream mediator of progesterone-induced lobuloalveolar maturation.[54] RANKL knockout mice show an almost identical mammary phenotype to PR knockout mice, including normal mammary ductal development but complete failure of the development of lobuloalveolar structures.[54]
Ductal development
Though to a far lesser extent than estrogen, which is the major mediator of mammary ductal development (via the ERα),[55][56] progesterone may be involved in ductal development of the mammary glands to some extent as well.[57] PR knockout mice or mice treated with the PR antagonist mifepristone show delayed although otherwise normal mammary ductal development at puberty.[57] In addition, mice modified to have overexpression of PRA display ductal hyperplasia,[54] and progesterone induces ductal growth in the mouse mammary gland.[57] Progesterone mediates ductal development mainly via induction of the expression of amphiregulin, the same growth factor that estrogen primarily induces the expression of to mediate ductal development.[57] These animal findings suggest that, while not essential for full mammary ductal development, progesterone seems to play a potentiating or accelerating role in estrogen-mediated mammary ductal development.[57]
Breast cancer risk
Progesterone also appears to be involved in the pathophysiology of breast cancer, though its role, and whether it is a promoter or inhibitor of breast cancer risk, has not been fully elucidated.[58] In any case, while most synthetic progestins, like medroxyprogesterone acetate, have been found to increase the risk of breast cancer in postmenopausal women in combination with estrogen as a component of hormone replacement therapy.[59][60] The combination may increase risk due to growth-promoting effects.[61] Whereas the combination of natural progesterone (or the atypical progestin dydrogesterone) with estrogen has not been found to increase breast cancer risk.[59][60]
Skin health
The estrogen receptor, as well as the progesterone receptor, have been detected in the skin, including in keratinocytes and fibroblasts.[62][63] At menopause and thereafter, decreased levels of female sex hormones result in atrophy, thinning, and increased wrinkling of the skin and a reduction in skin elasticity, firmness, and strength.[62][63] These skin changes constitute an acceleration in skin aging and are the result of decreased collagen content, irregularities in the morphology of epidermal skin cells, decreased ground substance between skin fibers, and reduced capillaries and blood flow.[62][63] The skin also becomes more dry during menopause, which is due to reduced skin hydration and surface lipids (sebum production).[62] Along with chronological aging and photoaging, estrogen deficiency in menopause is one of the three main factors that predominantly influences skin aging.[62]
Hormone replacement therapy, consisting of systemic treatment with estrogen alone or in combination with a progestogen, has well-documented and considerable beneficial effects on the skin of postmenopausal women.[62][63] These benefits include increased skin collagen content, skin thickness and elasticity, and skin hydration and surface lipids.[62][63] Topical estrogen has been found to have similar beneficial effects on the skin.[62] In addition, a study has found that topical 2% progesterone cream significantly increases skin elasticity and firmness and observably decreases wrinkles in peri- and postmenopausal women.[63] Skin hydration and surface lipids, on the other hand, did not significantly change with topical progesterone.[63] These findings suggest that progesterone, like estrogen, also has beneficial effects on the skin, and may be independently protective against skin aging.[63]
Libido
Progesterone and its neurosteroid active metabolite allopregnanolone appear to be importantly involved in libido in females.[64]
Homosexuality
Dr. Diana Fleischman, of the University of Portsmouth, and colleagues looked for a relationship between progesterone and sexual attitudes in 92 women. Their research, published in the Archives of Sexual Behavior found that women who had higher levels of progesterone scored higher on a questionnaire measuring homoerotic motivation. They also found that men who had high levels of progesterone were more likely to have higher homoerotic motivation scores after affiliative priming compared to men with low levels of progesterone.[65][66][67][68]
Nervous system
Progesterone, like pregnenolone and dehydroepiandrosterone (DHEA), belongs to an important group of endogenous steroids called neurosteroids. It can be metabolized within all parts of the central nervous system.[69]
Neurosteroids are neuromodulators, and are neuroprotective, neurogenic, and regulate neurotransmission and myelination.[70] The effects of progesterone as a neurosteroid are mediated predominantly through its interactions with non-nuclear PRs, namely the mPRs and PGRMC1, as well as certain other receptors, such as the σ1 and nACh receptors.
Brain damage
Previous studies have shown that progesterone supports the normal development of neurons in the brain, and that the hormone has a protective effect on damaged brain tissue. It has been observed in animal models that females have reduced susceptibility to traumatic brain injury and this protective effect has been hypothesized to be caused by increased circulating levels of estrogen and progesterone in females.[71]
Proposed mechanism
The mechanism of progesterone protective effects may be the reduction of inflammation that follows brain trauma and hemorrhage.[72][73]
Damage incurred by traumatic brain injury is believed to be caused in part by mass depolarization leading to excitotoxicity. One way in which progesterone helps to alleviate some of this excitotoxicity is by blocking the voltage-dependent calcium channels that trigger neurotransmitter release.[74] It does so by manipulating the signaling pathways of transcription factors involved in this release. Another method for reducing the excitotoxicity is by up-regulating the GABAA, a widespread inhibitory neurotransmitter receptor.[75]
Progesterone has also been shown to prevent apoptosis in neurons, a common consequence of brain injury.[76] It does so by inhibiting enzymes involved in the apoptosis pathway specifically concerning the mitochondria, such as activated caspase 3 and cytochrome c.
Not only does progesterone help prevent further damage, it has also been shown to aid in neuroregeneration.[77] One of the serious effects of traumatic brain injury includes edema. Animal studies show that progesterone treatment leads to a decrease in edema levels by increasing the concentration of macrophages and microglia sent to the injured tissue.[74][78] This was observed in the form of reduced leakage from the blood brain barrier in secondary recovery in progesterone treated rats. In addition, progesterone was observed to have antioxidant properties, reducing the concentration of oxygen free radicals faster than without.[75] There is also evidence that the addition of progesterone can also help remyelinate damaged axons due to trauma, restoring some lost neural signal conduction.[75] Another way progesterone aids in regeneration includes increasing the circulation of endothelial progenitor cells in the brain.[79] This helps new vasculature to grow around scar tissue which helps repair the area of insult.
Addiction
Progesterone enhances the function of serotonin receptors in the brain, so an excess or deficit of progesterone has the potential to result in significant neurochemical issues. This provides an explanation for why some people resort to substances that enhance serotonin activity such as nicotine, alcohol, and cannabis when their progesterone levels fall below optimal levels.[80]
- Sex differences in hormone levels may induce women to respond differently than men to nicotine. When women undergo cyclic changes or different hormonal transition phases (menopause, pregnancy, adolescence), there are changes in their progesterone levels.[81] Therefore, females have an increased biological vulnerability to nicotine's reinforcing effects compared to males and progesterone may be used to counter this enhanced vulnerability. This information supports the idea that progesterone can affect behavior.[80]
- Similar to nicotine, cocaine also increases the release of dopamine in the brain. The neurotransmitter is involved in the reward center and is one of the main neurotransmitters involved with substance abuse and reliance. In a study of cocaine users, it was reported that progesterone reduced craving and the feeling of being stimulated by cocaine. Thus, progesterone was suggested as an agent that decreases cocaine craving by reducing the dopaminergic properties of the drug.[82]
Societal
Among women, higher level of progesterone is correlated with lower level of competitiveness.[83]
Other effects
- Progesterone also has a role in skin elasticity and bone strength, in respiration, in nerve tissue and in female sexuality, and the presence of progesterone receptors in certain muscle and fat tissue may hint at a role in sexually dimorphic proportions of those.[84]
- During pregnancy, progesterone is said to decrease uterine irritability.[85]
- During pregnancy, progesterone helps to suppress immune responses of the mother to fetal antigens, which prevents rejection of the fetus.[85]
- Progesterone raises epidermal growth factor-1 (EGF-1) levels, a factor often used to induce proliferation, and used to sustain cultures, of stem cells.[86]
- Progesterone increases core temperature (thermogenic function) during ovulation.[87]
- Progesterone reduces spasm and relaxes smooth muscle. Bronchi are widened and mucus regulated. (PRs are widely present in submucosal tissue.)
- Progesterone acts as an antiinflammatory agent and regulates the immune response.
- Progesterone reduces gall-bladder activity.[88]
- Progesterone normalizes blood clotting and vascular tone, zinc and copper levels, cell oxygen levels, and use of fat stores for energy.
- Progesterone may affect gum health, increasing risk of gingivitis (gum inflammation).[89]
- Progesterone appears to prevent endometrial cancer (involving the uterine lining) by regulating the effects of estrogen.
- Progesterone plays an important role in the signaling of insulin release and pancreatic function, and may affect the susceptibility to diabetes or gestational diabetes.[90][91]
Biochemistry
Biosynthesis
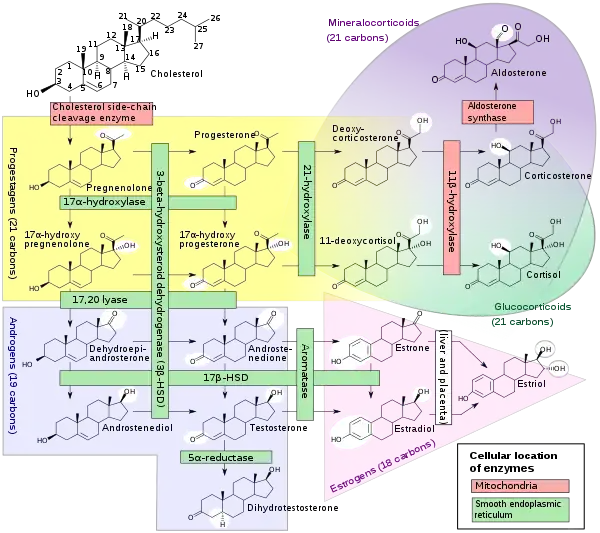
In mammals, progesterone, like all other steroid hormones, is synthesized from pregnenolone, which itself is derived from cholesterol.
Cholesterol undergoes double oxidation to produce 22R-hydroxycholesterol and then 20α,22R-dihydroxycholesterol. This vicinal diol is then further oxidized with loss of the side chain starting at position C22 to produce pregnenolone. This reaction is catalyzed by cytochrome P450scc.
The conversion of pregnenolone to progesterone takes place in two steps. First, the 3β-hydroxyl group is oxidized to a keto group and second, the double bond is moved to C4, from C5 through a keto/enol tautomerization reaction.[93] This reaction is catalyzed by 3β-hydroxysteroid dehydrogenase/δ5-4-isomerase.
Progesterone in turn is the precursor of the mineralocorticoid aldosterone, and after conversion to 17α-hydroxyprogesterone, of cortisol and androstenedione. Androstenedione can be converted to testosterone, estrone, and estradiol, highlighting the critical role of progesterone in testosterone synthesis.
Pregnenolone and progesterone can also be synthesized by yeast.[94]
During the luteal phase, approximately 25 mg of progesterone is secreted from the ovaries per day in women, while the adrenal glands produce about 1 mg of progesterone per day.[95]
| Sex | Sex hormone | Reproductive phase |
Blood production rate |
Gonadal secretion rate |
Metabolic clearance rate |
Reference range (serum levels) | |
|---|---|---|---|---|---|---|---|
| SI units | Non-SI units | ||||||
| Men | Androstenedione | – |
2.8 mg/day | 1.6 mg/day | 2200 L/day | 2.8–7.3 nmol/L | 80–210 ng/dL |
| Testosterone | – |
6.5 mg/day | 6.2 mg/day | 950 L/day | 6.9–34.7 nmol/L | 200–1000 ng/dL | |
| Estrone | – |
150 μg/day | 110 μg/day | 2050 L/day | 37–250 pmol/L | 10–70 pg/mL | |
| Estradiol | – |
60 μg/day | 50 μg/day | 1600 L/day | <37–210 pmol/L | 10–57 pg/mL | |
| Estrone sulfate | – |
80 μg/day | Insignificant | 167 L/day | 600–2500 pmol/L | 200–900 pg/mL | |
| Women | Androstenedione | – |
3.2 mg/day | 2.8 mg/day | 2000 L/day | 3.1–12.2 nmol/L | 89–350 ng/dL |
| Testosterone | – |
190 μg/day | 60 μg/day | 500 L/day | 0.7–2.8 nmol/L | 20–81 ng/dL | |
| Estrone | Follicular phase | 110 μg/day | 80 μg/day | 2200 L/day | 110–400 pmol/L | 30–110 pg/mL | |
| Luteal phase | 260 μg/day | 150 μg/day | 2200 L/day | 310–660 pmol/L | 80–180 pg/mL | ||
| Postmenopause | 40 μg/day | Insignificant | 1610 L/day | 22–230 pmol/L | 6–60 pg/mL | ||
| Estradiol | Follicular phase | 90 μg/day | 80 μg/day | 1200 L/day | <37–360 pmol/L | 10–98 pg/mL | |
| Luteal phase | 250 μg/day | 240 μg/day | 1200 L/day | 699–1250 pmol/L | 190–341 pg/mL | ||
| Postmenopause | 6 μg/day | Insignificant | 910 L/day | <37–140 pmol/L | 10–38 pg/mL | ||
| Estrone sulfate | Follicular phase | 100 μg/day | Insignificant | 146 L/day | 700–3600 pmol/L | 250–1300 pg/mL | |
| Luteal phase | 180 μg/day | Insignificant | 146 L/day | 1100–7300 pmol/L | 400–2600 pg/mL | ||
| Progesterone | Follicular phase | 2 mg/day | 1.7 mg/day | 2100 L/day | 0.3–3 nmol/L | 0.1–0.9 ng/mL | |
| Luteal phase | 25 mg/day | 24 mg/day | 2100 L/day | 19–45 nmol/L | 6–14 ng/mL | ||
Notes and sources
Notes: "The concentration of a steroid in the circulation is determined by the rate at which it is secreted from glands, the rate of metabolism of precursor or prehormones into the steroid, and the rate at which it is extracted by tissues and metabolized. The secretion rate of a steroid refers to the total secretion of the compound from a gland per unit time. Secretion rates have been assessed by sampling the venous effluent from a gland over time and subtracting out the arterial and peripheral venous hormone concentration. The metabolic clearance rate of a steroid is defined as the volume of blood that has been completely cleared of the hormone per unit time. The production rate of a steroid hormone refers to entry into the blood of the compound from all possible sources, including secretion from glands and conversion of prohormones into the steroid of interest. At steady state, the amount of hormone entering the blood from all sources will be equal to the rate at which it is being cleared (metabolic clearance rate) multiplied by blood concentration (production rate = metabolic clearance rate × concentration). If there is little contribution of prohormone metabolism to the circulating pool of steroid, then the production rate will approximate the secretion rate." Sources: See template. | |||||||
Distribution
Progesterone binds extensively to plasma proteins, including albumin (50–54%) and transcortin (43–48%).[96] It has similar affinity for albumin relative to the PR.[17]
Metabolism
The metabolism of progesterone is rapid and extensive and occurs mainly in the liver,[97][98][99] though enzymes that metabolize progesterone are also expressed widely in the brain, skin, and various other extrahepatic tissues.[69][100] Progesterone has an elimination half-life of only approximately 5 minutes in circulation.[97] The metabolism of progesterone is complex, and it may form as many as 35 different unconjugated metabolites when it is ingested orally.[99][101] Progesterone is highly susceptible to enzymatic reduction via reductases and hydroxysteroid dehydrogenases due to its double bond (between the C4 and C5 positions) and its two ketones (at the C3 and C20 positions).[99]
The major metabolic pathway of progesterone is reduction by 5α-reductase[69] and 5β-reductase into the dihydrogenated 5α-dihydroprogesterone and 5β-dihydroprogesterone, respectively.[98][99][102][103] This is followed by the further reduction of these metabolites via 3α-hydroxysteroid dehydrogenase and 3β-hydroxysteroid dehydrogenase into the tetrahydrogenated allopregnanolone, pregnanolone, isopregnanolone, and epipregnanolone.[104][98][99][102] Subsequently, 20α-hydroxysteroid dehydrogenase and 20β-hydroxysteroid dehydrogenase reduce these metabolites to form the corresponding hexahydrogenated pregnanediols (eight different isomers in total),[98][103] which are then conjugated via glucuronidation and/or sulfation, released from the liver into circulation, and excreted by the kidneys into the urine.[97][99] The major metabolite of progesterone in the urine is the 3α,5β,20α isomer of pregnanediol glucuronide, which has been found to constitute 15 to 30% of an injection of progesterone.[17][105] Other metabolites of progesterone formed by the enzymes in this pathway include 3α-dihydroprogesterone, 3β-dihydroprogesterone, 20α-dihydroprogesterone, and 20β-dihydroprogesterone, as well as various combination products of the enzymes aside from those already mentioned.[17][99][105][106] Progesterone can also first be hydroxylated (see below) and then reduced.[99] Endogenous progesterone is metabolized approximately 50% into 5α-dihydroprogesterone in the corpus luteum, 35% into 3β-dihydroprogesterone in the liver, and 10% into 20α-dihydroprogesterone.[107]
Relatively small portions of progesterone are hydroxylated via 17α-hydroxylase (CYP17A1) and 21-hydroxylase (CYP21A2) into 17α-hydroxyprogesterone and 11-deoxycorticosterone (21-hydroxyprogesterone), respectively,[101] and pregnanetriols are formed secondarily to 17α-hydroxylation.[108][109] Even smaller amounts of progesterone may be also hydroxylated via 11β-hydroxylase (CYP11B1) and to a lesser extent via aldosterone synthase (CYP11B2) into 11β-hydroxyprogesterone.[110][111] In addition, progesterone can be hydroxylated in the liver by other cytochrome P450 enzymes which are not steroid-specific.[112] 6β-Hydroxylation, which is catalyzed mainly by CYP3A4, is the major transformation, and is responsible for approximately 70% of cytochrome P450-mediated progesterone metabolism.[112] Other routes include 6α-, 16α-, and 16β-hydroxylation.[99] However, treatment of women with ketoconazole, a strong CYP3A4 inhibitor, had minimal effects on progesterone levels, producing only a slight and non-significant increase, and this suggests that cytochrome P450 enzymes play only a small role in progesterone metabolism.[113]
Metabolism of progesterone in humans[114]
|

Levels
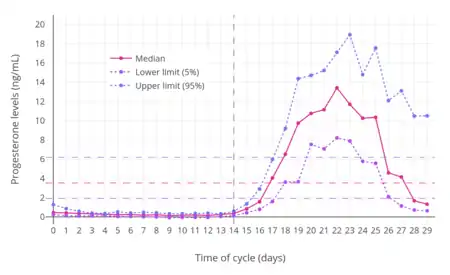
In women, progesterone levels are relatively low during the preovulatory phase of the menstrual cycle, rise after ovulation, and are elevated during the luteal phase, as shown in diagram below. Progesterone levels tend to be less than 2 ng/mL prior to ovulation, and greater than 5 ng/mL after ovulation. If pregnancy occurs, human chorionic gonadotropin is released maintaining the corpus luteum allowing it to maintain levels of progesterone. Between 7 and 9 weeks the placenta begins to produce progesterone in place of the corpus luteum, this process is named the luteal-placental shift.[116]
After the luteal-placental shift progesterone levels start to rise further and may reach 100 to 200 ng/mL at term. Whether a decrease in progesterone levels is critical for the initiation of labor has been argued and may be species-specific. After delivery of the placenta and during lactation, progesterone levels are very low.
Progesterone levels are low in children and postmenopausal women.[117] Adult males have levels similar to those in women during the follicular phase of the menstrual cycle.
Ranges
Blood test results should always be interpreted using the reference ranges provided by the laboratory that performed the results. Example reference ranges are listed below.
| Person type | Reference range for blood test | ||
|---|---|---|---|
| Lower limit | Upper limit | Unit | |
| Female - menstrual cycle | (see diagram below) | ||
| Female - postmenopausal | <0.2[118] | 1[118] | ng/mL |
| <0.6[119] | 3[119] | nmol/L | |
| Female on oral contraceptives | 0.34[118] | 0.92[118] | ng/mL |
| 1.1[119] | 2.9[119] | nmol/L | |
| Males ≥16 years | 0.27[118] | 0.9[118] | ng/mL |
| 0.86[119] | 2.9[119] | nmol/L | |
| Female or male 1–9 years | 0.1[118] | 4.1[118] or 4.5[118] | ng/mL |
| 0.3[119] | 13[119] | nmol/L | |
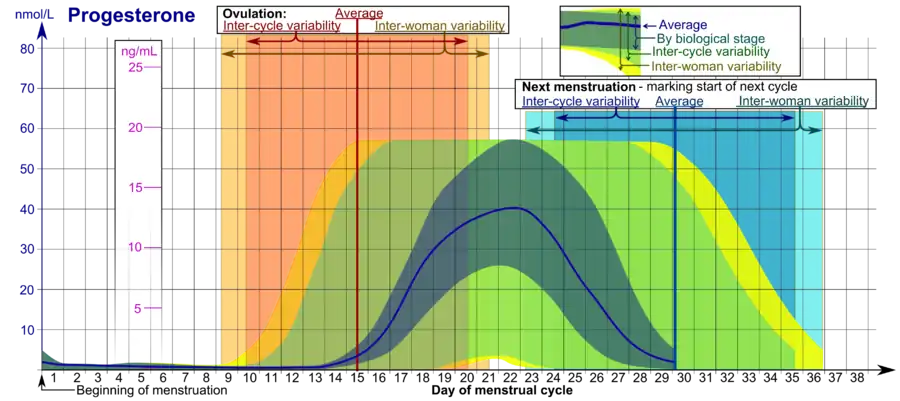
• The ranges denoted By biological stage may be used in closely monitored menstrual cycles in regard to other markers of its biological progression, with the time scale being compressed or stretched to how much faster or slower, respectively, the cycle progresses compared to an average cycle.
• The ranges denoted Inter-cycle variability are more appropriate to use in non-monitored cycles with only the beginning of menstruation known, but where the woman accurately knows her average cycle lengths and time of ovulation, and that they are somewhat averagely regular, with the time scale being compressed or stretched to how much a woman's average cycle length is shorter or longer, respectively, than the average of the population.
• The ranges denoted Inter-woman variability are more appropriate to use when the average cycle lengths and time of ovulation are unknown, but only the beginning of menstruation is given.
Animal
Progesterone is produced in high amounts in the ovaries (by the corpus luteum) from the onset of puberty to menopause, and is also produced in smaller amounts by the adrenal glands after the onset of adrenarche in both males and females. To a lesser extent, progesterone is produced in nervous tissue, especially in the brain, and in adipose (fat) tissue, as well.
During human pregnancy, progesterone is produced in increasingly high amounts by the ovaries and placenta. At first, the source is the corpus luteum that has been "rescued" by the presence of human chorionic gonadotropin (hCG) from the conceptus. However, after the 8th week, production of progesterone shifts to the placenta. The placenta utilizes maternal cholesterol as the initial substrate, and most of the produced progesterone enters the maternal circulation, but some is picked up by the fetal circulation and used as substrate for fetal corticosteroids. At term the placenta produces about 250 mg progesterone per day.
An additional animal source of progesterone is milk products. After consumption of milk products the level of bioavailable progesterone goes up.[121]
Plants
In at least one plant, Juglans regia, progesterone has been detected.[122] In addition, progesterone-like steroids are found in Dioscorea mexicana. Dioscorea mexicana is a plant that is part of the yam family native to Mexico.[123] It contains a steroid called diosgenin that is taken from the plant and is converted into progesterone.[124] Diosgenin and progesterone are also found in other Dioscorea species, as well as in other plants that are not closely related, such as fenugreek.
Another plant that contains substances readily convertible to progesterone is Dioscorea pseudojaponica native to Taiwan. Research has shown that the Taiwanese yam contains saponins — steroids that can be converted to diosgenin and thence to progesterone.[125]
Many other Dioscorea species of the yam family contain steroidal substances from which progesterone can be produced. Among the more notable of these are Dioscorea villosa and Dioscorea polygonoides. One study showed that the Dioscorea villosa contains 3.5% diosgenin.[126] Dioscorea polygonoides has been found to contain 2.64% diosgenin as shown by gas chromatography-mass spectrometry.[127] Many of the Dioscorea species that originate from the yam family grow in countries that have tropical and subtropical climates.[128]
Medical use
Progesterone is used as a medication. It is used in combination with estrogens mainly in hormone therapy for menopausal symptoms and low sex hormone levels in women.[101][129] It is also used in women to support pregnancy and fertility and to treat gynecological disorders.[130][131][132][133] Progesterone has been shown to prevent miscarriage in women with 1) vaginal bleeding early in their current pregnancy and 2) a previous history of miscarriage.[134] Progesterone can be taken by mouth, through the vagina, and by injection into muscle or fat, among other routes.[101]
Chemistry

Progesterone is a naturally occurring pregnane steroid and is also known as pregn-4-ene-3,20-dione.[135][136] It has a double bond (4-ene) between the C4 and C5 positions and two ketone groups (3,20-dione), one at the C3 position and the other at the C20 position.[135][136]
Semisynthesis 1
An economical semisynthesis of progesterone from the plant steroid diosgenin isolated from yams was developed by Russell Marker in 1940 for the Parke-Davis pharmaceutical company.[137] This synthesis is known as the Marker degradation. Additional semisyntheses of progesterone have also been reported starting from a variety of steroids. For the example, cortisone can be simultaneously deoxygenated at the C-17 and C-21 position by treatment with iodotrimethylsilane in chloroform to produce 11-keto-progesterone (ketogestin), which in turn can be reduced at position-11 to yield progesterone.[138]

Semisynthesis 2
Progesterone can also be made from the stigmasterol found in soybean oil also. c.f. Percy Julian.

Total synthesis

A total synthesis of progesterone was reported in 1971 by W.S. Johnson.[144] The synthesis begins with reacting the phosphonium salt 7 with phenyl lithium to produce the phosphonium ylide 8. The ylide 8 is reacted with an aldehyde to produce the alkene 9. The ketal protecting groups of 9 are hydrolyzed to produce the diketone 10, which in turn is cyclized to form the cyclopentenone 11. The ketone of 11 is reacted with methyl lithium to yield the tertiary alcohol 12, which in turn is treated with acid to produce the tertiary cation 13. The key step of the synthesis is the π-cation cyclization of 13 in which the B-, C-, and D-rings of the steroid are simultaneously formed to produce 14. This step resembles the cationic cyclization reaction used in the biosynthesis of steroids and hence is referred to as biomimetic. In the next step the enol orthoester is hydrolyzed to produce the ketone 15. The cyclopentene A-ring is then opened by oxidizing with ozone to produce 16. Finally, the diketone 17 undergoes an intramolecular aldol condensation by treating with aqueous potassium hydroxide to produce progesterone.[144]
History
The hormonal action of progesterone was discovered in 1929, following that of estrogen in 1923.[17][145][146] By 1931–1932, nearly pure crystalline material of high progestational activity had been isolated from the corpus luteum of animals, and by 1934, pure crystalline progesterone had been refined and obtained and the chemical structure of progesterone was determined.[17][145] This was achieved by Adolf Butenandt at the Chemisches Institut of Technical University in Danzig, who extracted this new compound from several thousand liters of urine.[147]
Chemical synthesis of progesterone from stigmasterol and pregnanediol was accomplished later that year.[145][148] Up to this point, progesterone, known generically as corpus luteum hormone, had been being referred to by several groups by different names, including corporin, lutein, luteosterone, and progestin.[17][149] In 1935, at the time of the Second International Conference on the Standardization of Sex Hormones in London, England, a compromise was made between the groups and the name progesterone (progestational steroidal ketone) was created.[17][150]
Veterinary use
The use of progesterone in tests dog breeding to pinpoint ovulation is becoming more widely used. There are several tests available but the most reliable test is a blood test with blood drawn by a veterinarian and sent to a lab for processing. Results can usually be obtained with 24 to 72 hours. The rationale for using progesterone tests is that increased numbers begin in close proximity to preovulatory surge in gonadotrophins and continue through ovulation and estrus. When progesterone levels reach certain levels they can signal the stage of estrus the female is. Prediction of birth date of the pending litter can be very accurate if ovulation date is known. Puppies deliver with a day or two of 9 weeks gestation in most cases. It is not possible to determine pregnancy using progesterone tests once a breeding has taken place however. This is due to the fact that, in dogs, progestrone levels remain elevated throughout the estrus period.[151]
References
- Jameson JL, De Groot LJ (25 February 2015). Endocrinology: Adult and Pediatric E-Book. Elsevier Health Sciences. p. 2179. ISBN 978-0-323-32195-2.
- Adler N, Pfaff D, Goy RW (6 Dec 2012). Handbook of Behavioral Neurobiology Volume 7 Reproduction (1st ed.). New York: Plenum Press. p. 189. ISBN 978-1-4684-4834-4. Retrieved 4 July 2015.
- "progesterone (CHEBI:17026)". ChEBI. European Molecular Biology Laboratory-EBI. Retrieved 4 July 2015.
- "Progesterone_msds".
- Stanczyk FZ (September 2002). "Pharmacokinetics and potency of progestins used for hormone replacement therapy and contraception". Reviews in Endocrine & Metabolic Disorders. 3 (3): 211–24. doi:10.1023/A:1020072325818. PMID 12215716. S2CID 27018468.
- Simon JA, Robinson DE, Andrews MC, Hildebrand JR, Rocci ML, Blake RE, Hodgen GD (July 1993). "The absorption of oral micronized progesterone: the effect of food, dose proportionality, and comparison with intramuscular progesterone". Fertility and Sterility. 60 (1): 26–33. doi:10.1016/S0015-0282(16)56031-2. PMID 8513955.
- Fritz MA, Speroff L (28 March 2012). Clinical Gynecologic Endocrinology and Infertility. Lippincott Williams & Wilkins. pp. 44–. ISBN 978-1-4511-4847-3.
- Marshall WJ, Marshall WJ, Bangert SK (2008). Clinical Chemistry. Elsevier Health Sciences. pp. 192–. ISBN 978-0-7234-3455-9.
- Yamazaki H, Shimada T (October 1997). "Progesterone and testosterone hydroxylation by cytochromes P450 2C19, 2C9, and 3A4 in human liver microsomes". Archives of Biochemistry and Biophysics. 346 (1): 161–9. doi:10.1006/abbi.1997.0302. PMID 9328296.
- McKay GA, Walters MR (6 February 2013). Lecture Notes: Clinical Pharmacology and Therapeutics. John Wiley & Sons. p. 33. ISBN 978-1-118-34489-7.
- Zutshi (1 January 2005). Hormones in Obstetrics and Gynaecology. Jaypee Brothers Publishers. p. 74. ISBN 978-81-8061-427-9.
- Cometti B (November 2015). "Pharmaceutical and clinical development of a novel progesterone formulation". Acta Obstetricia et Gynecologica Scandinavica. 94 (Suppl 161): 28–37. doi:10.1111/aogs.12765. PMID 26342177. S2CID 31974637.
- King TL, Brucker MC (25 October 2010). Pharmacology for Women's Health. Jones & Bartlett Publishers. pp. 372–373. ISBN 978-1-4496-5800-7.
- Baulieu E, Schumacher M (2000). "Progesterone as a neuroactive neurosteroid, with special reference to the effect of progesterone on myelination". Steroids. 65 (10–11): 605–12. doi:10.1016/s0039-128x(00)00173-2. PMID 11108866. S2CID 14952168.
- Prior JC (2019). "Progesterone Is Important for Transgender Women's Therapy — Applying Evidence for the Benefits of Progesterone in Ciswomen". The Journal of Clinical Endocrinology & Metabolism. 104 (4): 1181–1186. doi:10.1210/jc.2018-01777. PMID 30608551. S2CID 58620122.
Evidence has accrued that normal progesterone (and ovulation), as well as physiological estradiol levels, is necessary during ciswomen’s premenopausal menstrual cycles for current fertility and long-term health; transgender women may require progesterone therapy and similar potential physiological benefits
- Fischer J, Ganellin CR (2006). Analogue-based Drug Discovery. John Wiley & Sons. p. 47X. ISBN 9783527607495.
- Josimovich JB (11 November 2013). Gynecologic Endocrinology. Springer Science & Business Media. pp. 9, 25–29. ISBN 978-1-4613-2157-6.
- Taraborrelli S (November 2015). "Physiology, production and action of progesterone". Acta Obstetricia et Gynecologica Scandinavica. 94 Suppl 161: 8–16. doi:10.1111/aogs.12771. PMID 26358238. S2CID 40213725.
- Thomas P, Pang Y (2012). "Membrane progesterone receptors: evidence for neuroprotective, neurosteroid signaling and neuroendocrine functions in neuronal cells". Neuroendocrinology. 96 (2): 162–71. doi:10.1159/000339822. PMC 3489003. PMID 22687885.
- Valadez-Cosmes P, Vázquez-Martínez ER, Cerbón M, Camacho-Arroyo I (October 2016). "Membrane progesterone receptors in reproduction and cancer". Molecular and Cellular Endocrinology. 434: 166–75. doi:10.1016/j.mce.2016.06.027. PMID 27368976. S2CID 3826650.
- Meyer C, Schmid R, Schmieding K, Falkenstein E, Wehling M (February 1998). "Characterization of high affinity progesterone-binding membrane proteins by anti-peptide antiserum". Steroids. 63 (2): 111–6. doi:10.1016/s0039-128x(97)00143-8. PMID 9516722. S2CID 40096058.
- Kabe Y, Handa H, Suematsu M (July 2018). "Function and structural regulation of the carbon monoxide (CO)-responsive membrane protein PGRMC1". Journal of Clinical Biochemistry and Nutrition. 63 (1): 12–17. doi:10.3164/jcbn.17-132. PMC 6064819. PMID 30087538.
- Ryu CS, Klein K, Zanger UM (2017-03-27). "Membrane Associated Progesterone Receptors: Promiscuous Proteins with Pleiotropic Functions - Focus on Interactions with Cytochromes P450". Frontiers in Pharmacology. 8: 159. doi:10.3389/fphar.2017.00159. PMC 5366339. PMID 28396637.
- Maurice T, Urani A, Phan VL, Romieu P (November 2001). "The interaction between neuroactive steroids and the sigma1 receptor function: behavioral consequences and therapeutic opportunities". Brain Research. Brain Research Reviews. 37 (1–3): 116–32. doi:10.1016/s0165-0173(01)00112-6. PMID 11744080. S2CID 44931783.
- Johannessen M, Fontanilla D, Mavlyutov T, Ruoho AE, Jackson MB (February 2011). "Antagonist action of progesterone at σ-receptors in the modulation of voltage-gated sodium channels". American Journal of Physiology. Cell Physiology. 300 (2): C328-37. doi:10.1152/ajpcell.00383.2010. PMC 3043630. PMID 21084640.
- Rupprecht R, Reul JM, van Steensel B, Spengler D, Söder M, Berning B, et al. (October 1993). "Pharmacological and functional characterization of human mineralocorticoid and glucocorticoid receptor ligands". European Journal of Pharmacology. 247 (2): 145–54. doi:10.1016/0922-4106(93)90072-H. PMID 8282004.
- Elger W, Beier S, Pollow K, Garfield R, Shi SQ, Hillisch A (November 2003). "Conception and pharmacodynamic profile of drospirenone". Steroids. 68 (10–13): 891–905. doi:10.1016/j.steroids.2003.08.008. PMID 14667981. S2CID 41756726.
- Attardi BJ, Zeleznik A, Simhan H, Chiao JP, Mattison DR, Caritis SN (December 2007). "Comparison of progesterone and glucocorticoid receptor binding and stimulation of gene expression by progesterone, 17-alpha hydroxyprogesterone caproate, and related progestins". American Journal of Obstetrics and Gynecology. 197 (6): 599.e1–7. doi:10.1016/j.ajog.2007.05.024. PMC 2278032. PMID 18060946.
- Lei K, Chen L, Georgiou EX, Sooranna SR, Khanjani S, Brosens JJ, et al. (2012). "Progesterone acts via the nuclear glucocorticoid receptor to suppress IL-1β-induced COX-2 expression in human term myometrial cells". PLOS ONE. 7 (11): e50167. Bibcode:2012PLoSO...750167L. doi:10.1371/journal.pone.0050167. PMC 3509141. PMID 23209664.
- Paul SM, Purdy RH (March 1992). "Neuroactive steroids". FASEB Journal. 6 (6): 2311–22. doi:10.1096/fasebj.6.6.1347506. PMID 1347506. S2CID 221753076.
- Kliewer SA, Goodwin B, Willson TM (2002). "The nuclear pregnane X receptor: a key regulator of xenobiotic metabolism". Endocr. Rev. 23 (5): 687–702. doi:10.1210/er.2001-0038. PMID 12372848.
- Lehmann JM, McKee DD, Watson MA, Willson TM, Moore JT, Kliewer SA (1998). "The human orphan nuclear receptor PXR is activated by compounds that regulate CYP3A4 gene expression and cause drug interactions". J. Clin. Invest. 102 (5): 1016–23. doi:10.1172/JCI3703. PMC 508967. PMID 9727070.
- Meanwell NA (8 December 2014). Tactics in Contemporary Drug Design. Springer. pp. 161–. ISBN 978-3-642-55041-6.
- Legato MJ, Bilezikian JP (2004). Principles of Gender-specific Medicine. Gulf Professional Publishing. pp. 146–. ISBN 978-0-12-440906-4.
- Lemke TL, Williams DA (24 January 2012). Foye's Principles of Medicinal Chemistry. Lippincott Williams & Wilkins. p. 164. ISBN 978-1-60913-345-0.
- Estrogens—Advances in Research and Application: 2013 Edition: ScholarlyBrief. ScholarlyEditions. 21 June 2013. pp. 4–. ISBN 978-1-4816-7550-5.
- Strünker T, Goodwin N, Brenker C, Kashikar ND, Weyand I, Seifert R, Kaupp UB (March 2011). "The CatSper channel mediates progesterone-induced Ca2+ influx in human sperm". Nature. 471 (7338): 382–6. Bibcode:2011Natur.471..382S. doi:10.1038/nature09769. PMID 21412338. S2CID 4431334. Lay summary – Nature News.
- Lishko PV, Botchkina IL, Kirichok Y (March 2011). "Progesterone activates the principal Ca2+ channel of human sperm". Nature. 471 (7338): 387–91. Bibcode:2011Natur.471..387L. doi:10.1038/nature09767. PMID 21412339. S2CID 4340309.
- Kastner P, Krust A, Turcotte B, Stropp U, Tora L, Gronemeyer H, Chambon P (May 1990). "Two distinct estrogen-regulated promoters generate transcripts encoding the two functionally different human progesterone receptor forms A and B". The EMBO Journal. 9 (5): 1603–14. doi:10.1002/j.1460-2075.1990.tb08280.x. PMC 551856. PMID 2328727.
- Hallam SZ, Osuch JR (1 January 2006). Hormones and Breast Cancer in Post-Menopausal Women. IOS Press. p. 69. ISBN 978-1-58603-653-9.
- Johnson LR (2003). Essential Medical Physiology. Academic Press. p. 770. ISBN 978-0-12-387584-6.
- Coad J, Dunstall M (2011). Anatomy and Physiology for Midwives, with Pageburst online access,3: Anatomy and Physiology for Midwives. Elsevier Health Sciences. p. 413. ISBN 978-0-7020-3489-3.
- Landau RL, Bergenstal DM, Lugibihl K, Kascht ME (October 1955). "The metabolic effects of progesterone in man". The Journal of Clinical Endocrinology and Metabolism. 15 (10): 1194–215. doi:10.1210/jcem-15-10-1194. PMID 13263410.
- Correia JN, Conner SJ, Kirkman-Brown JC (May 2007). "Non-genomic steroid actions in human spermatozoa. "Persistent tickling from a laden environment"". Seminars in Reproductive Medicine. 25 (3): 208–19. doi:10.1055/s-2007-973433. PMID 17447210.
- Kirkman-Brown JC, Bray C, Stewart PM, Barratt CL, Publicover SJ (June 2000). "Biphasic elevation of [Ca(2+)](i) in individual human spermatozoa exposed to progesterone". Developmental Biology. 222 (2): 326–35. doi:10.1006/dbio.2000.9729. PMID 10837122.
- Kirkman-Brown JC, Barratt CL, Publicover SJ (March 2004). "Slow calcium oscillations in human spermatozoa". The Biochemical Journal. 378 (Pt 3): 827–32. doi:10.1042/BJ20031368. PMC 1223996. PMID 14606954.
- Harper CV, Barratt CL, Publicover SJ (October 2004). "Stimulation of human spermatozoa with progesterone gradients to simulate approach to the oocyte. Induction of [Ca(2+)](i) oscillations and cyclical transitions in flagellar beating". The Journal of Biological Chemistry. 279 (44): 46315–25. doi:10.1074/jbc.M401194200. PMID 15322137.
- Marieb, Elaine (2013). Anatomy & physiology. Benjamin-Cummings. p. 903. ISBN 9780321887603.
- Tosti E, Di Cosmo A, Cuomo A, Di Cristo C, Gragnaniello G (May 2001). "Progesterone induces activation in Octopus vulgaris spermatozoa". Molecular Reproduction and Development. 59 (1): 97–105. doi:10.1002/mrd.1011. PMID 11335951. S2CID 28390608.
- Bowen R (2000-08-06). "Placental Hormones". Retrieved 2008-03-12.
- Patel B, Elguero S, Thakore S, Dahoud W, Bedaiwy M, Mesiano S (2014). "Role of nuclear progesterone receptor isoforms in uterine pathophysiology". Human Reproduction Update. 21 (2): 155–73. doi:10.1093/humupd/dmu056. PMC 4366574. PMID 25406186.
- Di Renzo GC, Giardina I, Clerici G, Brillo E, Gerli S (July 2016). "Progesterone in normal and pathological pregnancy". Hormone Molecular Biology and Clinical Investigation. 27 (1): 35–48. doi:10.1515/hmbci-2016-0038. PMID 27662646. S2CID 32239449.
- Macias H, Hinck L (2012). "Mammary gland development". Wiley Interdisciplinary Reviews: Developmental Biology. 1 (4): 533–57. doi:10.1002/wdev.35. PMC 3404495. PMID 22844349.
- Hilton HN, Graham JD, Clarke CL (September 2015). "Minireview: Progesterone Regulation of Proliferation in the Normal Human Breast and in Breast Cancer: A Tale of Two Scenarios?". Molecular Endocrinology. 29 (9): 1230–42. doi:10.1210/me.2015-1152. PMC 5414684. PMID 26266959.
- Strauss JF, Barbieri RL (13 September 2013). Yen and Jaffe's Reproductive Endocrinology. Elsevier Health Sciences. pp. 236–. ISBN 978-1-4557-2758-2.
- Scaling AL, Prossnitz ER, Hathaway HJ (June 2014). "GPER mediates estrogen-induced signaling and proliferation in human breast epithelial cells and normal and malignant breast". Hormones & Cancer. 5 (3): 146–160. doi:10.1007/s12672-014-0174-1. PMC 4091989. PMID 24718936.
- Aupperlee MD, Leipprandt JR, Bennett JM, Schwartz RC, Haslam SZ (May 2013). "Amphiregulin mediates progesterone-induced mammary ductal development during puberty". Breast Cancer Research. 15 (3): R44. doi:10.1186/bcr3431. PMC 3738150. PMID 23705924.
- Kuhl H, Schneider HP (August 2013). "Progesterone--promoter or inhibitor of breast cancer". Climacteric. 16 Suppl 1: 54–68. doi:10.3109/13697137.2013.768806. PMID 23336704. S2CID 20808536.
- Fournier A, Berrino F, Clavel-Chapelon F (January 2008). "Unequal risks for breast cancer associated with different hormone replacement therapies: results from the E3N cohort study". Breast Cancer Research and Treatment. 107 (1): 103–11. doi:10.1007/s10549-007-9523-x. PMC 2211383. PMID 17333341.
- Campagnoli C, Clavel-Chapelon F, Kaaks R, Peris C, Berrino F (July 2005). "Progestins and progesterone in hormone replacement therapy and the risk of breast cancer". The Journal of Steroid Biochemistry and Molecular Biology. 96 (2): 95–108. doi:10.1016/j.jsbmb.2005.02.014. PMC 1974841. PMID 15908197.
- Asi N, Mohammed K, Haydour Q, Gionfriddo MR, Vargas OL, Prokop LJ, et al. (July 2016). "Progesterone vs. synthetic progestins and the risk of breast cancer: a systematic review and meta-analysis". Systematic Reviews. 5 (1): 121. doi:10.1186/s13643-016-0294-5. PMC 4960754. PMID 27456847.
- Raine-Fenning NJ, Brincat MP, Muscat-Baron Y (2003). "Skin aging and menopause : implications for treatment". American Journal of Clinical Dermatology. 4 (6): 371–8. doi:10.2165/00128071-200304060-00001. PMID 12762829. S2CID 20392538.
- Holzer G, Riegler E, Hönigsmann H, Farokhnia S, Schmidt JB, Schmidt B (September 2005). "Effects and side-effects of 2% progesterone cream on the skin of peri- and postmenopausal women: results from a double-blind, vehicle-controlled, randomized study". The British Journal of Dermatology. 153 (3): 626–34. doi:10.1111/j.1365-2133.2005.06685.x. PMID 16120154. S2CID 6077829.
- King SR (9 November 2012). Neurosteroids and the Nervous System. Springer Science & Business Media. pp. 44–46. ISBN 978-1-4614-5559-2.
- Fleischman DS, Fessler DM, Cholakians AE (July 2015). "Testing the Affiliation Hypothesis of Homoerotic Motivation in Humans: The Effects of Progesterone and Priming". Archives of Sexual Behavior. 44 (5): 1395–404. doi:10.1007/s10508-014-0436-6. PMID 25420899. S2CID 9864224.
- "Homosexuality may help us bond | UoP News".
- Having homosexual thoughts 'is an essential part of human evolution' study suggests The Telegraph
- Homosexuality May Have Evolved In Humans Because It Helps Us Bond, Scientists Say Huff Post
- Hanukoglu I, Karavolas HJ, Goy RW (April 1977). "Progesterone metabolism in the pineal, brain stem, thalamus and corpus callosum of the female rat". Brain Research. 125 (2): 313–24. doi:10.1016/0006-8993(77)90624-2. PMID 558037. S2CID 35814845.
- Schumacher M, Guennoun R, Robert F, Carelli C, Gago N, Ghoumari A, Gonzalez Deniselle MC, Gonzalez SL, Ibanez C, Labombarda F, Coirini H, Baulieu EE, De Nicola AF (June 2004). "Local synthesis and dual actions of progesterone in the nervous system: neuroprotection and myelination". Growth Hormone & IGF Research. 14 Suppl A: S18-33. doi:10.1016/j.ghir.2004.03.007. PMID 15135772.
- Roof RL, Hall ED (May 2000). "Gender differences in acute CNS trauma and stroke: neuroprotective effects of estrogen and progesterone". Journal of Neurotrauma. 17 (5): 367–88. doi:10.1089/neu.2000.17.367. PMID 10833057.
- Pan DS, Liu WG, Yang XF, Cao F (October 2007). "Inhibitory effect of progesterone on inflammatory factors after experimental traumatic brain injury". Biomedical and Environmental Sciences. 20 (5): 432–8. PMID 18188998.
- Jiang C, Zuo F, Wang Y, Wan J, Yang Z, Lu H, Chen W, Zang W, Yang Q, Wang J (June 2016). "Progesterone exerts neuroprotective effects and improves long-term neurologic outcome after intracerebral hemorrhage in middle-aged mice". Neurobiology of Aging. 42: 13–24. doi:10.1016/j.neurobiolaging.2016.02.029. PMC 4857017. PMID 27143417.
- Luoma JI, Stern CM, Mermelstein PG (August 2012). "Progesterone inhibition of neuronal calcium signaling underlies aspects of progesterone-mediated neuroprotection". The Journal of Steroid Biochemistry and Molecular Biology. 131 (1–2): 30–6. doi:10.1016/j.jsbmb.2011.11.002. PMC 3303940. PMID 22101209.
- Stein DG (March 2008). "Progesterone exerts neuroprotective effects after brain injury". Brain Research Reviews. 57 (2): 386–97. doi:10.1016/j.brainresrev.2007.06.012. PMC 2699575. PMID 17826842.
- Espinoza TR, Wright DW (2011). "The role of progesterone in traumatic brain injury". The Journal of Head Trauma Rehabilitation. 26 (6): 497–9. doi:10.1097/HTR.0b013e31823088fa. PMC 6025750. PMID 22088981.
- Jiang C, Zuo F, Wang Y, Lu H, Yang Q, Wang J (January 2017). "Progesterone Changes VEGF and BDNF Expression and Promotes Neurogenesis After Ischemic Stroke". Mol Neurobiol. 54 (1): 571–581. doi:10.1007/s12035-015-9651-y. PMC 4938789. PMID 26746666.
- Herson PS, Koerner IP, Hurn PD (May 2009). "Sex, sex steroids, and brain injury". Seminars in Reproductive Medicine. 27 (3): 229–39. doi:10.1055/s-0029-1216276. PMC 2675922. PMID 19401954.
- Li Z, Wang B, Kan Z, Zhang B, Yang Z, Chen J, Wang D, Wei H, Zhang JN, Jiang R (January 2012). "Progesterone increases circulating endothelial progenitor cells and induces neural regeneration after traumatic brain injury in aged rats". Journal of Neurotrauma. 29 (2): 343–53. doi:10.1089/neu.2011.1807. PMC 3261789. PMID 21534727.
- Lynch WJ, Sofuoglu M (December 2010). "Role of progesterone in nicotine addiction: evidence from initiation to relapse". Experimental and Clinical Psychopharmacology. 18 (6): 451–61. doi:10.1037/a0021265. PMC 3638762. PMID 21186920.
- Cosgrove KP, Esterlis I, McKee SA, Bois F, Seibyl JP, Mazure CM, Krishnan-Sarin S, Staley JK, Picciotto MR, O'Malley SS (April 2012). "Sex differences in availability of β2*-nicotinic acetylcholine receptors in recently abstinent tobacco smokers". Archives of General Psychiatry. 69 (4): 418–27. doi:10.1001/archgenpsychiatry.2011.1465. PMC 3508698. PMID 22474108.
- Mello NK, Knudson IM, Kelly M, Fivel PA, Mendelson JH (October 2011). "Effects of progesterone and testosterone on cocaine self-administration and cocaine discrimination by female rhesus monkeys". Neuropsychopharmacology. 36 (11): 2187–99. doi:10.1038/npp.2011.130. PMC 3176575. PMID 21796112.
- Buser, Thomas (2012-06-01). "The impact of the menstrual cycle and hormonal contraceptives on competitiveness". Journal of Economic Behavior & Organization. Gender Differences in Risk Aversion and Competition. 83 (1): 1–10. doi:10.1016/j.jebo.2011.06.006. ISSN 0167-2681.
- Sriram, D (2007). Medicinal Chemistry. New Delhi: Dorling Kindersley India Pvt. Ltd. p. 432. ISBN 978-81-317-0031-0.
- Blackburn S (14 April 2014). Maternal, Fetal, & Neonatal Physiology. Elsevier Health Sciences. pp. 92–. ISBN 978-0-323-29296-2.
- Faivre EJ, Lange CA (January 2007). "Progesterone receptors upregulate Wnt-1 to induce epidermal growth factor receptor transactivation and c-Src-dependent sustained activation of Erk1/2 mitogen-activated protein kinase in breast cancer cells". Molecular and Cellular Biology. 27 (2): 466–80. doi:10.1128/MCB.01539-06. PMC 1800800. PMID 17074804.
- Nosek, Thomas M. "Section 5/5ch9/s5ch9_13". Essentials of Human Physiology. Archived from the original on 2016-03-24.
- Hould FS, Fried GM, Fazekas AG, Tremblay S, Mersereau WA (December 1988). "Progesterone receptors regulate gallbladder motility". The Journal of Surgical Research. 45 (6): 505–12. doi:10.1016/0022-4804(88)90137-0. PMID 3184927.
- "Hormones and Oral Health". WebMD.
- Picard F, Wanatabe M, Schoonjans K, Lydon J, O'Malley BW, Auwerx J (November 2002). "Progesterone receptor knockout mice have an improved glucose homeostasis secondary to beta -cell proliferation". Proceedings of the National Academy of Sciences of the United States of America. 99 (24): 15644–8. doi:10.1073/pnas.202612199. PMC 137770. PMID 12438645.
- Brănişteanu DD, Mathieu C (March 2003). "Progesterone in gestational diabetes mellitus: guilty or not guilty?". Trends in Endocrinology and Metabolism. 14 (2): 54–6. doi:10.1016/S1043-2760(03)00003-1. PMID 12591170. S2CID 38209977.
- Häggström M, Richfield D (2014). "Diagram of the pathways of human steroidogenesis". WikiJournal of Medicine. 1 (1). doi:10.15347/wjm/2014.005. ISSN 2002-4436.
- Bewick PM (2002). Medicinal natural products: a biosynthetic approach. New York: Wiley. p. 244. ISBN 0-471-49641-3.
- Duport C, Spagnoli R, Degryse E, Pompon D (February 1998). "Self-sufficient biosynthesis of pregnenolone and progesterone in engineered yeast". Nature Biotechnology. 16 (2): 186–9. doi:10.1038/nbt0298-186. PMID 9487528. S2CID 852617.
- Lemke TL, Williams DA (24 January 2012). Foye's Principles of Medicinal Chemistry. Lippincott Williams & Wilkins. pp. 1397–. ISBN 978-1-60913-345-0.
- Progesterone - Drugs.com, retrieved 2015-08-23
- Falcone T, Hurd WW (2007). Clinical Reproductive Medicine and Surgery. Elsevier Health Sciences. pp. 22–. ISBN 978-0-323-03309-1.
- Cupps PT (20 February 1991). Reproduction in Domestic Animals. Elsevier. pp. 101–. ISBN 978-0-08-057109-6.
- Stanczyk FZ (November 2003). "All progestins are not created equal". Steroids. 68 (10–13): 879–90. doi:10.1016/j.steroids.2003.08.003. PMID 14667980. S2CID 44601264.
- Dowd FJ, Johnson B, Mariotti A (3 September 2016). Pharmacology and Therapeutics for Dentistry. Elsevier Health Sciences. pp. 448–. ISBN 978-0-323-44595-5.
- Kuhl H (August 2005). "Pharmacology of estrogens and progestogens: influence of different routes of administration". Climacteric. 8 Suppl 1: 3–63. doi:10.1080/13697130500148875. PMID 16112947. S2CID 24616324.
- Plant TM, Zeleznik AJ (15 November 2014). Knobil and Neill's Physiology of Reproduction. Academic Press. pp. 304–. ISBN 978-0-12-397769-4.
- Santoro NF, Neal-Perry G (11 September 2010). Amenorrhea: A Case-Based, Clinical Guide. Springer Science & Business Media. pp. 13–. ISBN 978-1-60327-864-5.
- Reddy DS (2010). "Neurosteroids: endogenous role in the human brain and therapeutic potentials". Progress in Brain Research. 186: 113–37. doi:10.1016/B978-0-444-53630-3.00008-7. PMC 3139029. PMID 21094889.
- Baulieu E, Kelly PA (30 November 1990). Hormones: From Molecules to Disease. Springer Science & Business Media. pp. 401–. ISBN 978-0-412-02791-8.
- Beranič N, Gobec S, Rižner TL (2011). "Progestins as inhibitors of the human 20-ketosteroid reductases, AKR1C1 and AKR1C3". Chem. Biol. Interact. 191 (1–3): 227–33. doi:10.1016/j.cbi.2010.12.012. PMID 21182831.
- Anderson GD, Odegard PS (October 2004). "Pharmacokinetics of estrogen and progesterone in chronic kidney disease". Adv Chronic Kidney Dis. 11 (4): 357–60. doi:10.1053/j.ackd.2004.07.001. PMID 15492972.
- Greenblatt JM, Brogan K (27 April 2016). Integrative Therapies for Depression: Redefining Models for Assessment, Treatment and Prevention. CRC Press. pp. 201–. ISBN 978-1-4987-0230-0.
- Graham C (2 December 2012). Reproductive Biology of the Great Apes: Comparative and Biomedical Perspectives. Elsevier. pp. 179–. ISBN 978-0-323-14971-6.
- Strushkevich, N.; Gilep, A. A.; Shen, L.; Arrowsmith, C. H.; Edwards, A. M.; Usanov, S. A.; Park, H. W. (2013). "Structural Insights into Aldosterone Synthase Substrate Specificity and Targeted Inhibition". Molecular Endocrinology (Baltimore, Md.). 27 (2): 315–324. doi:10.1210/me.2012-1287. PMC 5417327. PMID 23322723.
- van Rooyen D, Gent R, Barnard L, Swart AC (April 2018). "The in vitro metabolism of 11β-hydroxyprogesterone and 11-ketoprogesterone to 11-ketodihydrotestosterone in the backdoor pathway". The Journal of Steroid Biochemistry and Molecular Biology. 178: 203–212. doi:10.1016/j.jsbmb.2017.12.014. PMID 29277707. S2CID 3700135.
- de Azevedo Piccinato C (2008). Regulation of Steroid Metabolism and the Hepatic Transcriptome by Estradiol and Progesterone. pp. 24–25. ISBN 978-1-109-04632-8.
- Akalin S (January 1991). "Effects of ketoconazole in hirsute women". Acta Endocrinologica. 124 (1): 19–22. doi:10.1530/acta.0.1240019. PMID 1825737. S2CID 9831739.
- Aufrère MB, Benson H (June 1976). "Progesterone: an overview and recent advances". Journal of Pharmaceutical Sciences. 65 (6): 783–800. doi:10.1002/jps.2600650602. PMID 945344.
- Stricker R, Eberhart R, Chevailler MC, Quinn FA, Bischof P, Stricker R (2006). "Establishment of detailed reference values for luteinizing hormone, follicle stimulating hormone, estradiol, and progesterone during different phases of the menstrual cycle on the Abbott ARCHITECT analyzer". Clin. Chem. Lab. Med. 44 (7): 883–7. doi:10.1515/CCLM.2006.160. PMID 16776638. S2CID 524952.
- Csapo AI, Pulkkinen MU, Wiest WG (1973). "Effects of lutectomy and progestreone replacement therapy in early pregnancy patients". Am J Obstet Gynecol. 115 (6): 759–65. doi:10.1016/0002-9378(73)90517-6. PMID 4688578.
- NIH Clinical Center (2004-08-16). "Progesterone Historical Reference Ranges". United States National Institutes of Health. Archived from the original on 2009-01-09. Retrieved 2008-03-12.
- Progesterone Reference Ranges, Performed at the Clinical Center at the National Institutes of Health, Bethesda MD, 03Feb09
- Converted from mass values using molar mass of 314.46 g/mol
- Häggström, Mikael (2014). "Reference ranges for estradiol, progesterone, luteinizing hormone and follicle-stimulating hormone during the menstrual cycle". WikiJournal of Medicine. 1 (1). doi:10.15347/wjm/2014.001. ISSN 2002-4436.
- Goodson III WH, Handagama P, Moore II DH, Dairkee S (2007-12-13). "Milk products are a source of dietary progesterone". 30th Annual San Antonio Breast Cancer Symposium. pp. abstract # 2028. Retrieved 2008-03-12.
- Pauli GF, Friesen JB, Gödecke T, Farnsworth NR, Glodny B (March 2010). "Occurrence of progesterone and related animal steroids in two higher plants". Journal of Natural Products. 73 (3): 338–45. doi:10.1021/np9007415. PMID 20108949. S2CID 26467578.
- Applezweig N (May 1969). "Steroids". Chemical Week. 104: 57–72. PMID 12255132.
- Noguchi E, Fujiwara Y, Matsushita S, Ikeda T, Ono M, Nohara T (September 2006). "Metabolism of tomato steroidal glycosides in humans". Chemical & Pharmaceutical Bulletin. 54 (9): 1312–4. doi:10.1248/cpb.54.1312. PMID 16946542.
- Yang DJ, Lu TJ, Hwang LS (October 2003). "Isolation and identification of steroidal saponins in Taiwanese yam cultivar (Dioscorea pseudojaponica Yamamoto)". Journal of Agricultural and Food Chemistry. 51 (22): 6438–44. doi:10.1021/jf030390j. PMID 14558759.
- "Final report of the amended safety assessment of Dioscorea Villosa (Wild Yam) root extract". International Journal of Toxicology. 23 Suppl 2 (2_suppl): 49–54. 2004. doi:10.1080/10915810490499055. PMID 15513824. S2CID 962216.
- Niño J, Jiménez DA, Mosquera OM, Correa YM (2007). "Diosgenin quantification by HPLC in a Dioscorea polygonoides tuber collection from colombian flora". Journal of the Brazilian Chemical Society. 18 (5): 1073–1076. doi:10.1590/S0103-50532007000500030.
- Myoda T, Nagai T, Nagashima T (2005). Properties of starches in yam (Dioscorea spp.) tuber. Current Topics in Food Science and Technology. pp. 105–114. ISBN 81-308-0003-9.
- Wesp LM, Deutsch MB (2017). "Hormonal and Surgical Treatment Options for Transgender Women and Transfeminine Spectrum Persons". Psychiatr. Clin. North Am. 40 (1): 99–111. doi:10.1016/j.psc.2016.10.006. PMID 28159148.
- Ruan X, Mueck AO (November 2014). "Systemic progesterone therapy--oral, vaginal, injections and even transdermal?". Maturitas. 79 (3): 248–55. doi:10.1016/j.maturitas.2014.07.009. PMID 25113944.
- Filicori M (2015). "Clinical roles and applications of progesterone in reproductive medicine: an overview". Acta Obstet Gynecol Scand. 94 Suppl 161: 3–7. doi:10.1111/aogs.12791. PMID 26443945.
- Ciampaglia W, Cognigni GE (2015). "Clinical use of progesterone in infertility and assisted reproduction". Acta Obstet Gynecol Scand. 94 Suppl 161: 17–27. doi:10.1111/aogs.12770. PMID 26345161. S2CID 40753277.
- Choi SJ (2017). "Use of progesterone supplement therapy for prevention of preterm birth: review of literatures". Obstet Gynecol Sci. 60 (5): 405–420. doi:10.5468/ogs.2017.60.5.405. PMC 5621069. PMID 28989916.
- Coomarasamy A, Harb HM, Devall AJ, Cheed V, Roberts TE, Goranitis I, et al. (June 2020). "Progesterone to prevent miscarriage in women with early pregnancy bleeding: the PRISM RCT". Health Technology Assessment. 24 (33): 1–70. doi:10.3310/hta24330. PMC 7355406. PMID 32609084.
- Elks J (14 November 2014). The Dictionary of Drugs: Chemical Data: Chemical Data, Structures and Bibliographies. Springer. pp. 1024–. ISBN 978-1-4757-2085-3.
- Index Nominum 2000: International Drug Directory. Taylor & Francis. January 2000. pp. 880–. ISBN 978-3-88763-075-1.
- Marker RE, Krueger J (1940). "Sterols. CXII. Sapogenins. XLI. The Preparation of Trillin and its Conversion to Progesterone". J. Am. Chem. Soc. 62 (12): 3349–3350. doi:10.1021/ja01869a023.
- Numazawa M, Nagaoka M, Kunitama Y (September 1986). "Regiospecific deoxygenation of the dihydroxyacetone moiety at C-17 of corticoid steroids with iodotrimethylsilane". Chemical & Pharmaceutical Bulletin. 34 (9): 3722–6. doi:10.1248/cpb.34.3722. PMID 3815593.
- Heyl FW (1950). "Progesterone from 3-Acetoxybisnor-5-cholenaldehyde and 3-Ketobisnor-4-cholenaldehyde". Journal of the American Chemical Society. 72 (6): 2617–2619. doi:10.1021/ja01162a076.
- Slomp G (1958). "Ozonolysis. II. 1 The Effect of Pyridine on the Ozonolysis of 4,22-Stigmastadien-3-one 2". Journal of the American Chemical Society. 80 (4): 915–921. doi:10.1021/ja01537a041.
- Sundararaman P, Djerassi C (October 1977). "A convenient synthesis of progesterone from stigmasterol". The Journal of Organic Chemistry. 42 (22): 3633–4. doi:10.1021/jo00442a044. PMID 915584.
- "Nova Transcripts: Forgotten Genius". PBS.org. February 6, 2007.
- "Giants of the Past". lipidlibrary.aocs.org. Archived from the original on 2012-04-15.
- Johnson WS, Gravestock MB, McCarry BE (August 1971). "Acetylenic bond participation in biogenetic-like olefinic cyclizations. II. Synthesis of dl-progesterone". Journal of the American Chemical Society. 93 (17): 4332–4. doi:10.1021/ja00746a062. PMID 5131151.
- Coutinho EM, Segal SJ (1999). Is Menstruation Obsolete?. Oxford University Press. pp. 31–. ISBN 978-0-19-513021-8.
- Walker A (7 March 2008). The Menstrual Cycle. Routledge. pp. 49–. ISBN 978-1-134-71411-7.
- Piosik R (2003). "Adolf Butenandt und sein Wirken an der Technischen Hochschule Danzig". Chemkon. 10 (3): 135–138. doi:10.1002/ckon.200390038.
- Ginsburg B (6 December 2012). Premenstrual Syndrome: Ethical and Legal Implications in a Biomedical Perspective. Springer Science & Business Media. pp. 274–. ISBN 978-1-4684-5275-4.
- Rolleston HD (1936). The Endocrine Organs in Health and Disease: With an Historical Review. Oxford University Press, H. Milford. p. 406.
- Allen WM (October 1970). "Progesterone: how did the name originate?". Southern Medical Journal. 63 (10): 1151–5. doi:10.1097/00007611-197010000-00012. PMID 4922128. S2CID 35867375.
- Refsal K (February 2009). "Interpretation of Serum Progesterone Results for Management of Breeding in Dogs" (PDF). Webcd.endo.ref.
External links
- Progesterone MS Spectrum
- Progesterone at the US National Library of Medicine Medical Subject Headings (MeSH)
- Kimball JW (2007-05-27). "Progesterone". Kimball's Biology Pages. Archived from the original on 2008-06-18. Retrieved 2008-06-18.