SbtB protein
SbtB, which stands for sodium-bicarbonate-transporter B, is a protein found in bacteria. This small soluble protein has been classified as a new member of the P-II family that is involved in signal transduction. This protein has been demonstrated to participate in numerous processes including carbon sensing mechanisms in cyanobacteria.
| Membrane-associated protein slr1513 | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Identifiers | |||||||
| Organism | |||||||
| Symbol | slr1513 | ||||||
| Alt. symbols | SbtB | ||||||
| UniProt | P73954 | ||||||
| |||||||
Carbon concentrating mechanisms
Currently, most of the oxygen on planet earth derives from oxygenic photosynthesis.[3] Some phototrophic prokaryotes such as cyanobacteria developed the ability to carry out oxygenic photosynthesis around 2.7 billion years ago. Moreover, 2 billion years ago planet earth was struck with "The Great Oxygenation event" also known as the oxygen crisis. Rising levels of atmospheric O2, mainly due to oxygenic photosynthesis carried out by cyanobacteria nearly caused the mass death of anaerobic organisms. Having to face the evolutionary pressure of dropping ambient CO2 levels, cyanobacteria coped by evolving carbon concentrating mechanisms (CCM). Thus, these carbon concentrating mechanisms, also known as "DIC-pumps",[4] are found in most photosynthetic microorganisms, such as unicellular green algea and cyanobacteria. These environmental adaptations vastly improve photosynthetic performance and survival. Indeed, this is achieved by accumulating intracellular inorganic carbon (Ci) providing elevated CO2 levels around the primary CO2 fixing enzyme, Rubisco.
Although, until very recently it was poorly understood how these photoprotic procaryotes could sense the fluctuations in inorganic carbon (Ci), scientists have recently discovered that a gene, SbtB, encoding a small soluble protein (SbtB) which may have an important role in the process. SbtB participates in sensing fluctuations of CO2 concentrations in the environment and therefore adjusts CO2 fixation in according to its surroundings.
This discovery has been groundbreaking as earth and all of its inhabitants depend on photosynthetic carbon dioxide fixation to form organic carbon. In addition, cyanobacteria are some of the oldest organisms on earth and any additional knowledge about how they operate internally offers insight on the earliest forms of life on earth. These procaryotic organisms may enclose the answers to some of the most challenging questions ever posed in regards to the origin of life as we know it.
Structure
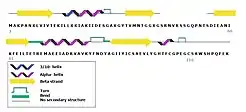
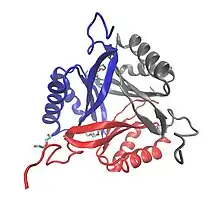
'SbtB' is a homo trimeric protein. The monomer consists of 120 amino acid residues with molecular weight of 11875 Da.[2] In addition, the monomer contains the following secondary structures:
- Beta-sheets (34% of the chain): there is a total of 4 strands composed by a total of 41 residues.
- Helical structures (26% of the chain): 4 helical structures. The first one consisting of a 3/10-helix and an alpha helix. The rest are exclusively made up by alpha helix structures.
- 3 T-turns. The flexible T loop is mostly disordered which is common in the PII like proteins[2]
- the beta sheets and alpha-helical structures are connected by loops.
A crystallographic structure of SbtB protein has been obtained from ScSbtB (Synechocystis sp.[2]) The apo-ScSbtB shows a canonical ferredoxin-like fold in every subunit. As seen before, the molecular structure of the three subunits is identical. At the N terminus the first beta-sheets face the C terminus. The carboxyl terminus of the last cysteine forms a hydrogen bond with the N terminus. This interaction allows the stabilization of N and C terminus.[2]
Ligand binding
ScSbtB has shown to bind ATP, ADP and cAMP[2] with dissociation constants (Kd) of 46, 19, and 11 μM respectively.[2] The ring opened form of cAMP, AMP binds with much lower affinity.
Gene
This protein is coded by the 'SbtB' gene,[5] also called 'CMM_2535' gene. Assembled in the genome of the clavibacter michiganensis specie,[6] It is a putative serine protease gene that belongs to the PTHR10795 family, which contains 23545 species.
It is assembled in the genome of the species Clavibacter michiganensis.[6] This species is part of the family Microbacteriaceae, and has five subspecies. The subspecies that contains this gene in its genome is C. m. subsp. michiganensis. The latter is involved in bacterial wilt and canker of tomato.[7]
It's located in the circular chromosome[8] of this kind of microorganisms, which contains 2984 coding genes and 3,297,891 bit/s (base pairs). Specifically, sbtB is located between the 2,851,656-2,855,336 bit/s and is composed by six different motifs[9] which are shown below.
Gene motifs
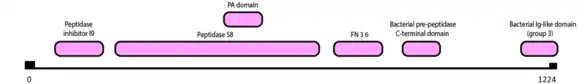
In terms of the aminoacid sequence (1224 aminoacids), the sbtB gene has the following motifs:
| Motif | From | To | E value |
|---|---|---|---|
| Peptidase Inhibitor I9 | 85 | 184 | 1.4e-11 |
| Peptidase S8 (subtilase family) | 213 | 694 | 6.3e-42 |
| PA domain | 485 | 559 | 1.5e-09 |
| FN 3 6 | 735 | 828 | 5.9e-23 |
| Bacterial pre-peptidase C-terminal domain | 895 | 961 | 0.002 |
| Bacterial Ig-like domain (group 3) | 1146 | 1224 | 1.6e-11 |
Chromosome statistics
The circular chromosome of this bacteria specie has the following characteristics:
| Length (bps) | 3,297,891 |
|---|---|
| Coding genes | 2,984 |
| Non coding genes | 106 |
| Pseudogenes | 24 |
Transcript gene: sbtB-1
It has a transcript gene called 'sbtB-1'.[10] This specific transcript gene has 1 coding exon and is annotated with 23 domains. In term of statistics, it has a transcript length of 3681 bit/s and a translation length of 1226 residues. It is located between the 2,855,336 - 2,851,656 base pairs.
Function
Serine-type endopeptidase activity
SbtB is part of the S8A family[12] inside the SB Clan[13] as classified by the peptidise database MEROPS[14][15] as well as uniprot.[16] This family is characterised by their main function:[17][18] the hydrolysis of peptide bonds.[18] This is possible due to the catalytic triad in their active site of their protein sequence: Aspartate, Serine and Histidine (the triad is presented in this specific order in the S8 family). The serine nucleophile responsible for the cleave of peptide bonds is activated by the contribution of the acidic residue (Asp) and the basic residue (His).
Carbon concentration
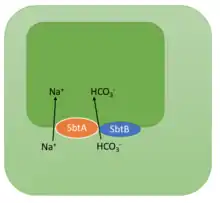
SbtB is part of the cyanobacterial CCM (CO2-concentrating mechanism) system.[11] It acts as a post-translational regulator (inhibitor) of the SbtA protein (one of the three sodium-dependant bicarbonate (Na+/HCO3−) symporters in the cytoplasmic membrane) by binding to it during the dark hours to prevent rising levels of Na+ from growing and becoming toxic for the cyanobacterial cell.[19] The SbtB protein is crucial for the photosynthetic process as it inhibits the SbtbA protein via direct interaction between both proteins. It is therefore a regulator of the Na+/HCO3− transporter[11] and is responsible for the cyanobacterial inorganic carbon response via cAMP sensing.[2]
Importance in binding processes
SbtB not only takes part in the binding process of ATP and ADP,[2] but is also the first ever known protein to bind the nucleotide cAMP (Cyclic adenosine monophosphate). This cyclic nucleotide is a single-phosphate nucleotide with a cyclic bond between the phosphate and the sugar, and is a derivative from ATP. Furthermore, cAMP is a key-signaling molecule in a wide range of processes including the state of carbon metabolism in all organisms. Until now cAMP was widely known for its important role in the maintenance of glucose balance, but in identifying SbtB researchers have discovered a new carbon sensing mechanism through cAMP.[2] SbtB's role is gaining importance as research shows it is a direct participer in the sensing of inorganic carbon fluctuation in cyanobacteria.[2]
References
- Humphrey W, Dalke A, Schulten K (February 1996). "VMD: visual molecular dynamics". Journal of Molecular Graphics. 14 (1): 33–8, 27–8. doi:10.1016/0263-7855(96)00018-5. PMID 8744570.
- PDB: 5O3S; Selim KA, Haase F, Hartmann MD, Hagemann M, Forchhammer K (May 2018). "II-like signaling protein SbtB links cAMP sensing with cyanobacterial inorganic carbon response". Proceedings of the National Academy of Sciences of the United States of America. 115 (21): E4861–E4869. doi:10.1073/pnas.1803790115. PMC 6003466. PMID 29735650. Lay summary – Phys.org.
- Cummins EP, Selfridge AC, Sporn PH, Sznajder JI, Taylor CT (March 2014). "Carbon dioxide-sensing in organisms and its implications for human disease". Cellular and Molecular Life Sciences. 71 (5): 831–45. doi:10.1007/s00018-013-1470-6. PMC 3945669. PMID 24045706.
- Ghoshal D, Goyal A (December 2000). "Carbon concentration mechanisms in photosynthetic microorganisms" (PDF). Indian Journal of Biochemistry & Biophysics. 37 (6): 383–94. PMID 11355625.
- "Clavibacter michiganensis subsp. michiganensis NCPPB 382". Ensembl Genomes Bacteria.
- "Clavibacter michiganensis subsp. michiganensis". KEGG Genome.
- Eichenlaub R, Gartemann KH (2011). "The Clavibacter michiganensis subspecies: molecular investigation of gram-positive bacterial plant pathogens". Annual Review of Phytopathology. 49: 445–64. doi:10.1146/annurev-phyto-072910-095258. PMID 21438679.
- "Chromosome: 2,851,656-2,855,336 - Region in detail - Clavibacter michiganensis subsp. michiganensis NCPPB 382". Ensembl Genomes Bacteria. Retrieved 2018-10-19.
- "SSDB Motif Search Result: cmi:CMM_2535". KEGG GENES Database.
- "Transcript: sbtB-1 CAN02617". Ensembl Bacteria.
- Du J, Förster B, Rourke L, Howitt SM, Price GD (December 2014). "Characterisation of cyanobacterial bicarbonate transporters in E. coli shows that SbtA homologs are functional in this heterologous expression system". PLOS ONE. 9 (12): e115905. Bibcode:2014PLoSO...9k5905D. doi:10.1371/journal.pone.0115905. PMC 4275256. PMID 25536191.
- "Family S8". MEROPS - the Peptidase Database. Retrieved 2018-10-17.
- "Summary for clan SB". MEROPS - the Peptidase Database. Retrieved 2018-10-17.
- "MEROPS - the Peptidase Database". www.ebi.ac.uk. Retrieved 2018-10-17.
- "Index of Peptidase Gene Names". MEROPS - the Peptidase Database.
- Universal protein resource accession number A5CU31 for "sbtB - Putative serine protease, peptidase family S8A - Clavibacter michiganensis subsp. michiganensis (strain NCPPB 382) - sbtB gene & protein" at UniProt.
- "AmiGO 2: Term Details for "serine-type endopeptidase activity" (GO:0004252)". Amigo. Gene Ontology Consortium. Retrieved 2018-10-17.
- "GO:0008236 serine-type peptidase activity". QuickGO. EMBL-EBI. Retrieved 2018-10-17.
- Wey L (2016). McKay D (ed.). "Regulation of sodium-dependent bicarbonate transporter, SbtA" (PDF). The ANU Undergraduate Research Journal. 7 (2015). doi:10.22459/AURJ.07.2015.11.