Cohesin
Cohesin is a protein complex that mediates sister chromatid cohesion, homologous recombination and DNA looping. Cohesin is formed of SMC3, SMC1, SCC1 and SCC3 (SA1 or SA2 in humans). Cohesin holds sister chromatids together after DNA replication until anaphase when removal of cohesin leads to separation of sister chromatids. The complex forms a ring-like structure and it is believed that sister chromatids are held together by entrapment inside the cohesin ring. Cohesin is a member of the SMC family of protein complexes which includes Condensin, MukBEF and SMC-ScpAB.

Cohesin was separately discovered in budding yeast by Douglas Koshland[1] and Kim Nasmyth.[2]
Structure
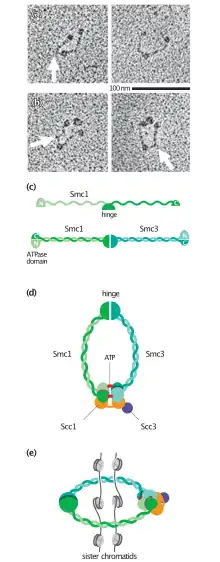
Cohesin is a multi-subunit protein complex, made up of SMC1, SMC3, RAD21 and SCC3 (SA1 or SA2).[3] SMC1 and SMC3 are members of the Structural Maintenance of Chromosomes (SMC) family. SMC proteins have two main structural characteristics: an ATP-binding cassette-like 'head' domain with ATPase activity (formed by the interaction of the N- and C- terminals) and a hinge domain that allows dimerization of SMCs. The head and the hinge domains are connected to each other via long anti-parallel coiled coils. The dimer is present in a V-shaped form, connected by the hinges.
The N-terminal domain of RAD21 contains two α-helices which forms a three helix bundle with the coiled coil of SMC3.[4] The central region of RAD21 is thought to be largely unstructured but contains several binding sites for regulators of cohesin. This includes a binding site for SA1 or SA2,[5] recognition motifs for separase cleavage[6] and a region that is competitively bound by PDS5A, PDS5B or NIPBL.[7][8][9] The C-terminal domain of RAD21 forms a winged helix that binds two β-sheets in the Smc1 head domain.[10]
Once RAD21 binds the SMC proteins, SCC3 can also associate with RAD21. When RAD21 binds on both SMC1 and SMC3, the cohesin complex forms a closed ring structure. The interfaces between the SMC subunits and RAD21 can open to allow DNA to pass in and out of the cohesin ring.[11]
While structures are available for many of the subunits and their interfaces, a structure of the entire cohesin complex has not been solved. Our knowledge of the conformation of cohesin comes largely from electron microscopy. These studies have revealed cohesin in numerous conformations including rings, elongated rods and most recently in a folded conformations. It is not known which conformation is predominant inside the cell and whether some are induced by sample preparation.[12]
Function
The cohesin ring has many functions:
1. It is used to keep the sister chromatids connected with each other during metaphase ensuring that during mitosis (and meiosis), each sister chromatid segregates to opposite poles. Without cohesin, the cell would be unable to control sister chromatid segregation since there would be no way of ensuring whether the spindle fiber attached on each sister chromatid is from a different pole.
2. It facilitates spindle attachment onto chromosomes.
3. It facilitates DNA repair by recombination.
4. Recently many novel functions of cohesin have been discovered in many different cellular processes. Cohesin has been shown to be responsible for transcription regulation, DNA double strand break repair, chromosome condensation, pairing of homologous chromosomes during meiosis I, mono-orientation of sister kinetochores during meiosis I, non-homologous centromere coupling, chromosome architecture and rearrangement, DNA replication etc.[13]
Dissociation of sister chromatid cohesion
The anaphase promoting complex associated to Cdc20 (APC/C-cdc20) marks Securin (anaphase inhibitor) for degradation by the proteasome. Securin is cleaved at anaphase, following APC/C-cdc20 mediated degradation, and it renders separase (a protease, inhibited by the association with securin) to cleave the kleisin subunit. An alpha-kleisin is associated with the cohesin complex, linking both SMC 3 and SMC 1 together, with the exact kleisin varying between mitosis and meiosis (Scc1 and Rec8 respectively), and its cleavage ultimately leads to the removal of cohesin from chromosomes.[14]
Dissociation of sister chromatids cohesion defines anaphase onset, which establishes two sets of identical chromosomes at each pole of the cell (telophase). Then the two daughter cells separate, and a new round of the cell cycle freshly starts in each one, at the stage of G0. When cells are ready to divide, because cell size is big enough or because they receive the appropriate stimulus,[15] they activate the mechanism to enter into the G1 stage of cell cycle, and they duplicate most organelles during S (synthesis) phase, including their centrosome. Therefore, when the cell division process will end, each daughter cell will receive a complete set of organelles. At the same time, during S phase all cells must duplicate their DNA very precisely, a process termed DNA replication. Once DNA replication has finished, in eukaryotes the DNA molecule is compacted and condensed, to form the mitotic chromosomes, each one constituted by two sister chromatids, which stay held together by the establishment of cohesion between them; each chromatid is a complete DNA molecule, attached via microtubules to one of the two centrosomes of the dividing cell, located at opposed poles of the cell. To avoid premature sister chromatid separation, the APC/C is maintained in an inactive state bound to different molecules, which are part of a complex mechanism termed the spindle assembly checkpoint.
Mechanism of Sister Chromatid Cohesion
It is not clear how the cohesin ring links sister chromatids together. There are two possible scenarios:
- Cohesin subunits bind to each sister chromatid and form a bridge between the two.
- Since cohesin has a ring structure, it is able to encircle both sister chromatids.
Current evidence suggests that the second scenario is the most likely. Proteins that are essential for sister chromatid cohesion, such as Smc3 and Scc1, do not regulate the formation of covalent bonds between cohesin and DNA, indicating that DNA interaction is not sufficient for cohesion.[11] In addition, disturbing the ring structure of cohesin through cleavage of Smc3 or Scc1 triggers premature sister chromatid segregation in vivo.[16] This shows that the ring structure is important for cohesin’s function.
Early studies suggested various ways in which cohesin may entrap DNA,[17] including as a monomer that holds both homologues together, and a "hand-cuff" model where two intertwining cohesin complexes each hold one sister chromatid. While some studies support the idea of a hand-cuff model,[17] the model is inconsistent with a number of experimental observations,[18] and is generally considered to entrap chromatin as a monomer.
Even though the ring hypothesis appears to be valid, there are still questions about the number of rings required to hold sister chromatids together. One possibility is that one ring surrounds the two chromatids. Another possibility involves the creation of a dimer where each ring surrounds one sister chromatid. The two rings are connected to each other through formation of a bridge that holds the two sister chromatids together.
The topology and structure of these subunits has been best characterized in budding yeast,[19][20] but the sequence conservation of these proteins and biochemical and electron microscopic observations imply that cohesin complexes in other species are very similar in their structure, .
The cohesin complex is established during the initial stages of S-phase. The complexes associate with chromosomes before DNA replication occurs. Once cells start replicating their DNA, cohesin rings close and link the sister chromatids together.[11] Cohesin complexes must be present during S-phase in order for cohesion to take place. It is unclear, however, how cohesin is loaded on the chromosomes during G1. There are two proposed hypotheses so far:
- The ATPase domain of the SMC proteins interacts with DNA and this interaction initially mediates the loading of cohesin complexes on chromosomes.
- Several proteins aid in the loading process. For example, Scc2 and Scc4 are both required for cohesin to load in budding yeast.
Localization of cohesin rings
Cohesin binding along the chromosomal DNA is considered to be dynamic and its location changes based on gene transcription, specific DNA sequence and presence of chromosome-associated proteins. There are three possible scenarios:
- Cohesin location is influenced by the orientation of neighboring genes and it is most frequently located in areas of convergent transcription. Gene orientation depends on the direction of transcription and can be of three types: head-to-head, head-to-tail and tail-to-tail. The tail-to-tail configuration results in the convergence of transcription machinery. One hypothesis states that the RNA polymerase “pushes” cohesin along the DNA, causing them to move towards the direction of the RNA polymerases. Changing the transcription pattern of genes changes the location of cohesin indicating that localization of cohesin may depend on transcription.[21]
- In another model, chromatin loop extrusion is pushed by transcription generated supercoiling ensuring also that cohesin relocalizes quickly and loops grow with reasonable speed and in a good direction. In addition, the supercoiling-driven loop extrusion mechanism is consistent with earlier explanations proposing why topologically associating domains (TADs) flanked by convergent CTCF binding sites form more stable chromatin loops than TADs flanked by divergent CTCF binding sites. In this model, the supercoiling also stimulates enhancer promoter contacts and it is proposed that transcription of eRNA sends the first wave of supercoiling that can activate mRNA transcription in a given TAD.[22]
- A few cohesin rings are found in chromosome arms that have AT-rich DNA sequences indicating that DNA sequence may be an independent factor of cohesin binding.[21]
- Cohesin rings, especially in budding yeast, are also located in the region surrounding the centromere.[21] Two hypotheses may explain this: the presence of repetitive heterochromatic DNA in centromeres and the presence of chromosome-associated proteins. For example, Schizosaccharomyces pombe have multiple copies of specific heterochromatic DNA whose involvement in cohesion binding has been proven. Budding yeast lacks repetitive sequences and, therefore, requires a different mechanism for cohesion binding. Evidence suggests that binding of cohesin to the budding yeast centromere region depends on chromosome-associated proteins of the kinetochore that mediate cohesion association to pericentric regions (the kinetochore is an enhancer of pericentric cohesin binding).[23]
Cohesin and CTCF
Many chromatin loops are formed by so called loop extrusion mechanism, when the cohesin ring moves actively along the two DNA double helices, translocating one of them with respect to the other. Thus, the loop can become smaller or larger. The loop extrusion process stops when cohesin encounters the architectural chromatin protein CTCF. The CTCF site needs to be in a proper orientation to stop cohesin.
Meiosis
Cohesin proteins SMC1ß, SMC3, REC8 and STAG3 appear to participate in cohesion of sister chromatids throughout the meiotic process in human oocytes.[24] SMC1ß, REC8 and STAG3 proteins are meiosis specific cohesins.
The STAG3 protein appears to be essential for female meiosis. A homozygous frameshift mutation in the Stag3 gene was identified in a large consanguineous family with premature ovarian failure.[25] Also, female mice deficient in STAG3 are sterile, and their fetal oocytes arrest at early prophase 1.
Evolution
Cohesin structure and function has been conserved in evolution. The SMC proteins are found in prokaryotes and have been conserved through evolution. The coils of SMC1 and SMC3 are conserved with an amino acid divergence of less than 0.5%.[26]
| Name | Saccharomyces cerevisiae | Schizosaccharomyces pombe | Drosophila | Vertebrates |
|---|---|---|---|---|
| Smc1 | Smc1 | Psm1 | DmSmc1 | Smc1 |
| Smc3 | Smc3 | Psm3 | DmSmc3 | Smc3 |
| Scc1 | Mcd1/Pds3 | Rad21 | DmRad21 | Rad21 |
| Scc3 | Scc3 | Psc3 | DmSA | SA1 and SA2 |
Clinical significance
The term "cohesinopathy" has been used to describe conditions affecting the cohesin complex.[27][28][29]
These conditions include:
- Cornelia de Lange Syndrome
- Roberts syndrome
- Warsaw breakage syndrome
- Many types of malignancies
References
- Guacci, V; Koshland, D; Strunnikov, A (3 October 1997). "A direct link between sister chromatid cohesion and chromosome condensation revealed through the analysis of MCD1 in S. cerevisiae". Cell. 91 (1): 47–57. doi:10.1016/s0092-8674(01)80008-8. PMC 2670185. PMID 9335334.
- Michaelis, C; Ciosk, R; Nasmyth, K (3 October 1997). "Cohesins: chromosomal proteins that prevent premature separation of sister chromatids". Cell. 91 (1): 35–45. doi:10.1016/s0092-8674(01)80007-6. PMID 9335333. S2CID 18572651.
- Losada A, Hirano M, Hirano T (1998). "Identification of Xenopus SMC protein complexes required for sister chromatid cohesion". Genes Dev. 12 (13): 1986–1997. doi:10.1101/gad.12.13.1986. PMC 316973. PMID 9649503.CS1 maint: multiple names: authors list (link)
- Gligoris, TG; Scheinost, JC; Bürmann, F; Petela, N; Chan, KL; Uluocak, P; Beckouët, F; Gruber, S; Nasmyth, K; Löwe, J (21 November 2014). "Closing the cohesin ring: structure and function of its Smc3-kleisin interface". Science. 346 (6212): 963–7. Bibcode:2014Sci...346..963G. doi:10.1126/science.1256917. PMC 4300515. PMID 25414305.
- Hara, K; Zheng, G; Qu, Q; Liu, H; Ouyang, Z; Chen, Z; Tomchick, DR; Yu, H (October 2014). "Structure of cohesin subcomplex pinpoints direct shugoshin-Wapl antagonism in centromeric cohesion". Nature Structural & Molecular Biology. 21 (10): 864–70. doi:10.1038/nsmb.2880. PMC 4190070. PMID 25173175.
- Uhlmann, F; Lottspeich, F; Nasmyth, K (1 July 1999). "Sister-chromatid separation at anaphase onset is promoted by cleavage of the cohesin subunit Scc1". Nature. 400 (6739): 37–42. Bibcode:1999Natur.400...37U. doi:10.1038/21831. PMID 10403247. S2CID 4354549.
- Petela, NJ; Gligoris, TG; Metson, J; Lee, BG; Voulgaris, M; Hu, B; Kikuchi, S; Chapard, C; Chen, W; Rajendra, E; Srinivisan, M; Yu, H; Löwe, J; Nasmyth, KA (21 June 2018). "Scc2 Is a Potent Activator of Cohesin's ATPase that Promotes Loading by Binding Scc1 without Pds5". Molecular Cell. 70 (6): 1134–1148.e7. doi:10.1016/j.molcel.2018.05.022. PMC 6028919. PMID 29932904.
- Kikuchi, S; Borek, DM; Otwinowski, Z; Tomchick, DR; Yu, H (1 November 2016). "Crystal structure of the cohesin loader Scc2 and insight into cohesinopathy". Proceedings of the National Academy of Sciences of the United States of America. 113 (44): 12444–12449. doi:10.1073/pnas.1611333113. PMC 5098657. PMID 27791135.
- Muir, KW; Kschonsak, M; Li, Y; Metz, J; Haering, CH; Panne, D (8 March 2016). "Structure of the Pds5-Scc1 Complex and Implications for Cohesin Function". Cell Reports. 14 (9): 2116–2126. doi:10.1016/j.celrep.2016.01.078. PMID 26923589.
- Haering, CH; Schoffnegger, D; Nishino, T; Helmhart, W; Nasmyth, K; Löwe, J (24 September 2004). "Structure and stability of cohesin's Smc1-kleisin interaction". Molecular Cell. 15 (6): 951–64. doi:10.1016/j.molcel.2004.08.030. PMID 15383284.
- Gruber S, Haering CH, Nasmyth K (March 2003). "Chromosomal cohesin forms a ring". Cell. 112 (6): 765–77. doi:10.1016/S0092-8674(03)00162-4. PMID 12654244.
- Yatskevich, S; Rhodes, J; Nasmyth, K (3 December 2019). "Organization of Chromosomal DNA by SMC Complexes". Annual Review of Genetics. 53: 445–482. doi:10.1146/annurev-genet-112618-043633. PMID 31577909.
- Mehta GD, Kumar R, Srivastava S, Ghosh SK (August 2013). "Cohesin: functions beyond sister chromatid cohesion". FEBS Letters. 587 (15): 2299–312. doi:10.1016/j.febslet.2013.06.035. PMID 23831059. S2CID 39397443.
- Mehta GD, Rizvi SM, Ghosh SK (August 2012). "Cohesin: a guardian of genome integrity". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1823 (8): 1324–42. doi:10.1016/j.bbamcr.2012.05.027. PMID 22677545.
- Conlon I, Raff M (January 1999). "Size control in animal development". Cell. 96 (2): 235–44. doi:10.1016/S0092-8674(00)80563-2. PMID 9988218.
- Peters JM, Tedeschi A, Schmitz J (November 2008). "The cohesin complex and its roles in chromosome biology". Genes & Development. 22 (22): 3089–114. doi:10.1101/gad.1724308. PMID 19056890.
- Zhang N, Kuznetsov SG, Sharan SK, Li K, Rao PH, Pati D (December 2008). "A handcuff model for the cohesin complex". The Journal of Cell Biology. 183 (6): 1019–31. doi:10.1083/jcb.200801157. PMC 2600748. PMID 19075111.
- Nasmyth K (October 2011). "Cohesin: a catenase with separate entry and exit gates?". Nature Cell Biology. 13 (10): 1170–7. doi:10.1038/ncb2349. PMID 21968990. S2CID 25382204.
- Haering, CH; Löwe, J; Hochwagen, A; Nasmyth, K (April 2002). "Molecular architecture of SMC proteins and the yeast cohesin complex". Molecular Cell. 9 (4): 773–88. doi:10.1016/s1097-2765(02)00515-4. PMID 11983169.
- Haering, CH; Farcas, AM; Arumugam, P; Metson, J; Nasmyth, K (17 July 2008). "The cohesin ring concatenates sister DNA molecules" (PDF). Nature. 454 (7202): 297–301. Bibcode:2008Natur.454..297H. doi:10.1038/nature07098. PMID 18596691. S2CID 1190883.
- Ross KE, Cohen-Fix O (July 2004). "Molecular biology: cohesins slip sliding away". Nature. 430 (6999): 520–1. Bibcode:2004Natur.430..520R. doi:10.1038/430520b. PMID 15282594. S2CID 52818523.
- Racko D, Benedetti F, Dorier J, Stasiak A (13 November 2017). "Transcription-induced supercoiling as the driving force of chromatin loop extrusion during formation of TADs in interphase chromosomes". Nucleic Acids Res. 46 (4): 1648–1660. doi:10.1093/nar/gkx1123. PMC 5829651. PMID 29140466.
- Weber SA, Gerton JL, Polancic JE, DeRisi JL, Koshland D, Megee PC (September 2004). "The kinetochore is an enhancer of pericentric cohesin binding". PLOS Biology. 2 (9): E260. doi:10.1371/journal.pbio.0020260. PMC 490027. PMID 15309047.
- Garcia-Cruz R, Brieño MA, Roig I, Grossmann M, Velilla E, Pujol A, Cabero L, Pessarrodona A, Barbero JL, Garcia Caldés M (September 2010). "Dynamics of cohesin proteins REC8, STAG3, SMC1 beta and SMC3 are consistent with a role in sister chromatid cohesion during meiosis in human oocytes". Human Reproduction. 25 (9): 2316–27. doi:10.1093/humrep/deq180. PMID 20634189.
- Caburet S, Arboleda VA, Llano E, Overbeek PA, Barbero JL, Oka K, Harrison W, Vaiman D, Ben-Neriah Z, García-Tuñón I, Fellous M, Pendás AM, Veitia RA, Vilain E (March 2014). "Mutant cohesin in premature ovarian failure". The New England Journal of Medicine. 370 (10): 943–949. doi:10.1056/NEJMoa1309635. PMC 4068824. PMID 24597867.
- White GE, Erickson HP (2009). "The coiled coils of cohesin are conserved in animals, but not in yeast". PLOS ONE. 4 (3): e4674. Bibcode:2009PLoSO...4.4674W. doi:10.1371/journal.pone.0004674. PMC 2650401. PMID 19262687.
- Gard S, Light W, Xiong B, Bose T, McNairn AJ, Harris B, Fleharty B, Seidel C, Brickner JH, Gerton JL (November 2009). "Cohesinopathy mutations disrupt the subnuclear organization of chromatin". The Journal of Cell Biology. 187 (4): 455–62. doi:10.1083/jcb.200906075. PMC 2779225. PMID 19948494.
- van der Lelij P, Chrzanowska KH, Godthelp BC, Rooimans MA, Oostra AB, Stumm M, Zdzienicka MZ, Joenje H, de Winter JP (February 2010). "Warsaw breakage syndrome, a cohesinopathy associated with mutations in the XPD helicase family member DDX11/ChlR1". American Journal of Human Genetics. 86 (2): 262–6. doi:10.1016/j.ajhg.2010.01.008. PMC 2820174. PMID 20137776.
- van der Lelij P, Godthelp BC, van Zon W, van Gosliga D, Oostra AB, Steltenpool J, de Groot J, Scheper RJ, Wolthuis RM, Waisfisz Q, Darroudi F, Joenje H, de Winter JP (September 2009). Warburton PE (ed.). "The cellular phenotype of Roberts syndrome fibroblasts as revealed by ectopic expression of ESCO2". PLOS ONE. 4 (9): e6936. Bibcode:2009PLoSO...4.6936V. doi:10.1371/journal.pone.0006936. PMC 2734174. PMID 19738907.
Further reading
- Mehta GD, Rizvi SM, Ghosh SK (August 2012). "Cohesin: a guardian of genome integrity". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1823 (8): 1324–42. doi:10.1016/j.bbamcr.2012.05.027. PMID 22677545.
- Mehta GD, Kumar R, Srivastava S, Ghosh SK (August 2013). "Cohesin: functions beyond sister chromatid cohesion". FEBS Letters. 587 (15): 2299–312. doi:10.1016/j.febslet.2013.06.035. PMID 23831059. S2CID 39397443.
- Michaelis C, Ciosk R, Nasmyth K (October 1997). "Cohesins: chromosomal proteins that prevent premature separation of sister chromatids". Cell. 91 (1): 35–45. doi:10.1016/S0092-8674(01)80007-6. PMID 9335333.
- Guacci V, Koshland D, Strunnikov A (October 1997). "A direct link between sister chromatid cohesion and chromosome condensation revealed through the analysis of MCD1 in S. cerevisiae". Cell. 91 (1): 47–57. doi:10.1016/S0092-8674(01)80008-8. PMC 2670185. PMID 9335334.
- Tóth A, Ciosk R, Uhlmann F, Galova M, Schleiffer A, Nasmyth K (February 1999). "Yeast cohesin complex requires a conserved protein, Eco1p(Ctf7), to establish cohesion between sister chromatids during DNA replication". Genes & Development. 13 (3): 320–33. doi:10.1101/gad.13.3.320. PMC 316435. PMID 9990856.
- Uhlmann F, Lottspeich F, Nasmyth K (July 1999). "Sister-chromatid separation at anaphase onset is promoted by cleavage of the cohesin subunit Scc1". Nature. 400 (6739): 37–42. Bibcode:1999Natur.400...37U. doi:10.1038/21831. PMID 10403247. S2CID 4354549.
External links
 Media related to Cohesins at Wikimedia Commons
Media related to Cohesins at Wikimedia Commons- cohesin at the US National Library of Medicine Medical Subject Headings (MeSH)
