Epiboly
Epiboly describes one of the five major types of cell movements that occur in the Gastrulation stage of embryonic development of some organisms.[1] Epibolic movement is the way in which a layer epithelial cells spreads. This can be achieved in multiple ways.
When undergoing epiboly, a monolayer of cells must undergo a physical change in shape in order to spread. Alternatively, multiple layers of cells can also undergo epiboly as the position of cells is changed or the cell layers undergo intercalation. While human embryos do not experience epiboly, this movement can be studied in sea urchins, tunicates, amphibians, and most commonly zebrafish.
Zebrafish
General movements
Epiboly in zebrafish is the first coordinated cell movement, and begins once the embryo has completed the blastula stage.[2] At this point the zebrafish embryo contains three portions: an epithelial monolayer known as the enveloping layer (EVL), a yolk syncytial layer (YSL) which is a membrane-enclosed group of nuclei that lie on top of the yolk cell, and the deep cells (DEL) of the blastoderm which will eventually form the embryo's three germ layers (ectoderm, mesoderm, and endoderm). The EVL, YSL, and DEL all undergo epiboly.

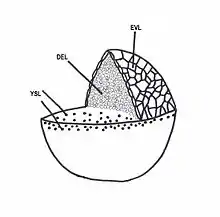
Radial intercalation occurs in the DEL. Interior cells of the blastoderm move towards the outer cells, thus "intercalating" with each other. The blastoderm begins to thin as it spreads toward the vegetal pole of the embryo until it has completely engulfed the yolk cell.[3] The EVL also moves vegetally during epiboly, increasing its surface area as it spreads. Work in the ray-finned fish Fundulus has shown that no large rearrangements occur in the EVL; instead, cells at the leading edge of the EVL align and constrict.[4][5] The YSL also moves towards the vegetal pole, spreading along the surface of the yolk and migrating slightly ahead of the blastomeres.[6] Once epiboly is complete, the DEL, EVL, and YSL have engulfed the yolk cell, forming a closure known as the blastopore.
Cytoskeletal and cell adhesion components
Completion of epiboly requires the coordination of cytoskeletal changes across the embryo. The YSL appears to play a prominent role in this process. Studies on Fundulus demonstrated that the YSL is capable of undergoing epiboly even when the blastoderm has been removed, however, the blastoderm cannot undergo epiboly in the absence of the YSL.[7] In zebrafish, there is a microtubule array in the yolk that extends from the animal to the vegetal pole of the embryo, and that contracts as epiboly progresses.[8] Treating embryos with the microtubule depolymerizing agent nocodazole completely blocks epiboly of the YSL and partially blocks epiboly of the blastoderm, while treating with the microtubule stabilizing agent taxol blocks epiboly of all cell layers.[8] There is also evidence for the importance of actin-based structures in epiboly. Ring-like structures of filamentous actin have been observed at the leading edge of the enveloping layer, where it contacts the yolk cell.[9] It is thought that a network of filamentous actin in the yolk might constrict in a myosin-II dependent manner to close the blastopore at the end of epiboly, via a "purse-string mechanism".[10] Treating embryos with the actin destabilizer cytochalasin b results in delayed or arrested epiboly.[9]
There is still debate on the extent to which the DEL and EVL epibolic movements are active movements.[11] The EVL contacts the YSL by means of tight junctions. It is thought that these contacts allow the YSL to "tow" the EVL towards the vegetal pole.[7] Claudin E is a molecule found in tight junctions that appears to be expressed in the EVL and required for normal zebrafish epiboly, supporting this hypothesis.[12] Additionally, zebrafish embryos that fail to make a fully differentiated EVL show defects in epibolic movements of the DEL, EVL, and YSL, suggesting a requirement for a normal EVL for the epiboly of all three cell layers.[13]
The cell-cell adhesion molecule E-cadherin has been shown to be required for the radial intercalation of the deep cells.[3] Many other molecules involved in cell-cell contact are implicated in zebrafish epiboly, including G alpha (12/13) which interacts with E-Cadherin and actin, as well as the cell adhesion molecule EpCam in the EVL, which may modulate adhesion with the underlying deep cells.[14][15]
Other vertebrates
Epibolic movements have been conserved in vertebrates. Though most work on epiboly has been done in fish, there is also a body of work concerning epiboly in the African clawed frog, Xenopus laevis. Comparisons of epiboly in amniotes, teleosts and X. laevis show that the key movement of epiboly in the fish and frog is radial intercalation while in amniotes it would appear to be cell division in the plane of the epithelium. All groups undergo cell shape changes such as the characteristic flattening of cells to increase surface area.[22]
References
- Developmental Biology, 10e. Sinauer Associates, Inc. 2014. Table 5.2.
- Warga RM, Kimmel CB (April 1990). "Cell movements during epiboly and gastrulation in zebrafish". Development. 108 (4): 569–80. PMID 2387236.
- Donald A. Kane; Karen N. McFarland; Rachel M. Warga (2005-03-01). "Mutations in half baked/E-cadherin block cell behaviors that are necessary for teleost epiboly". Development. 132 (5): 1105–16. doi:10.1242/dev.01668. PMID 15689372.
- Kimmel CB, Warga RM (November 1987). "Indeterminate cell lineage of the zebrafish embryo". Dev. Biol. 124 (1): 269–80. doi:10.1016/0012-1606(87)90478-7. PMID 3666309.
- C.B. Kimmel; R.M. Warga; T.F. Schilling (1990-04-01). "Origin and organization of the zebrafish fate map". Development. 108 (4): 581–94. PMID 2387237.
- D'Amico LA, Cooper MS (December 2001). "Morphogenetic domains in the yolk syncytial layer of axiating zebrafish embryos". Dev. Dyn. 222 (4): 611–24. doi:10.1002/dvdy.1216. PMID 11748830. Archived from the original on 2013-01-05.
- Betchaku T, Trinkaus JP (December 1978). "Contact relations, surface activity, and cortical microfilaments of marginal cells of the enveloping layer and of the yolk syncytial and yolk cytoplasmic layers of fundulus before and during epiboly". J. Exp. Zool. 206 (3): 381–426. doi:10.1002/jez.1402060310. PMID 568653.
- L. Solnica-Krezel; W. Driever (1994-09-01). "Microtubule arrays of the zebrafish yolk cell: organization and function during epiboly". Development. 120 (9): 2443–55. PMID 7956824.
- Cheng JC, Miller AL, Webb SE (October 2004). "Organization and function of microfilaments during late epiboly in zebrafish embryos". Dev. Dyn. 231 (2): 313–23. doi:10.1002/dvdy.20144. PMID 15366008.
- Mathias Köppen; Beatriz García Fernández; Lara Carvalho; Antonio Jacinto; Carl-Philipp Heisenberg (2006-07-15). "Coordinated cell-shape changes control epithelial movement in zebrafish and Drosophila". Development. 133 (14): 2671–81. doi:10.1242/dev.02439. PMID 16794032.
- A. Bruce and R. Winklbauer 03-P005 Zebra fish epiboly as a model of vertebrate cell rearrangement, Mechanisms of Development 126 (2009)
- Siddiqui M, Sheikh H, Tran C, Bruce A (2010). "The tight junction component claudin E is required for zebra fish epiboly". Developmental Dynamics. 239 (2): 715–722. doi:10.1002/dvdy.22172. PMID 20014098.
- Fukazawa C, Santiago C, Park K, Deery W, Gomez de la Torre Canny S, Holterhoff C, Wagner DS (October 2010). "poky/chuk/ikk1 is required for differentiation of the zebrafish embryonic epidermis". Developmental Biology. 346 (2): 272–83. doi:10.1016/j.ydbio.2010.07.037. PMC 2956273. PMID 20692251.
- Fang Lin; Songhai Chen; Diane S. Sepich; Jennifer Ray Panizzi; Sherry G. Clendenon; James A. Marrs; Heidi E. Hamm; Solnica-Krezel, L. (2009-03-23). "Gα12/13 regulate epiboly by inhibiting E-cadherin activity and modulating the actin cytoskeleton". The Journal of Cell Biology. 184 (6): 909–21. doi:10.1083/jcb.200805148. PMC 2664974. PMID 19307601.
- Slanchev K; Carney TJ; Stemmler MP; et al. (July 2009). Mullins, Mary C (ed.). "The Epithelial Cell Adhesion Molecule EpCAM Is Required for Epithelial Morphogenesis and Integrity during Zebrafish Epiboly and Skin Development". PLoS Genet. 5 (7): e1000563. doi:10.1371/journal.pgen.1000563. PMC 2700972. PMID 19609345.
- Mungo Marsden; Douglas W. DeSimone (2001-09-15). "Regulation of cell polarity, radial intercalation and epiboly in Xenopus: novel roles for integrin and fibronectin". Development. 128 (18): 3635–47. PMID 11566866.
- M. Hammerschmidt; F. Pelegri; M.C. Mullins; D.A. Kane; M. Brand; F.J. van Eeden; M. Furutani-Seiki; Granato, M; Haffter, P (1996-12-01). "Mutations affecting morphogenesis during gastrulation and tail formation in the zebrafish, Danio rerio". Development. 123 (1): 143–51. PMID 9007236.
- Martina Nagel; Emilios Tahinci; Karen Symes; Rudolf Winklbauer (2004-06-01). "Guidance of mesoderm cell migration in the Xenopus gastrula requires PDGF signaling". Development. 131 (11): 2727–36. doi:10.1242/dev.01141. PMID 15128658.
- Oates AC; Lackmann M; Power MA; et al. (May 1999). "An early developmental role for eph-ephrin interaction during vertebrate gastrulation". Mech. Dev. 83 (1–2): 77–94. doi:10.1016/S0925-4773(99)00036-2. PMID 10381569.
- Conway G, Margoliath A, Wong-Madden S, Roberts RJ, Gilbert W (April 1997). "Jak1 kinase is required for cell migrations and anterior specification in zebrafish embryos". Proc. Natl. Acad. Sci. U.S.A. 94 (7): 3082–7. doi:10.1073/pnas.94.7.3082. PMC 20325. PMID 9096349.
- Holloway BA, Gomez de la Torre Canny S, Ye Y, Slusarski DC, Freisinger CM, Dosch R, Chou MM, Wagner DS, Mullins MC (March 2009). Barsh GS (ed.). "A Novel Role for MAPKAPK2 in Morphogenesis during Zebrafish Development". PLoS Genetics. 5 (3): e1000413. doi:10.1371/journal.pgen.1000413. PMC 2652113. PMID 19282986.
- Solnica-Krezel L (March 2005). "Conserved patterns of cell movements during vertebrate gastrulation". Curr. Biol. 15 (6): R213–28. doi:10.1016/j.cub.2005.03.016. PMID 15797016.
