Mutation bias
Mutation bias refers to a pattern in which some type of mutation occurs more often than expected under uniformity. The types are most often defined by the molecular nature of the mutational change (see examples below), but sometimes they are based on downstream effects, e.g., Ostrow, et al.[1] refer to the tendency for mutations to increase body size in nematodes as a mutation bias.
Scientific context
The concept of mutation bias appears in several scientific contexts, most commonly in molecular studies of evolution, where mutation biases may be invoked to account for such phenomena as systematic differences in codon usage or genome composition between species.[2] The short tandem repeat (STR) loci used in forensic identification may show biased patterns of gain and loss of repeats.[3] In cancer research, some types of tumors have distinctive mutational signatures that reflect differences in the contributions of mutational pathways. Mutational signatures have proved useful in both detection and treatment.
Recent studies of the emergence of resistance to anti-microbials and anti-cancer drugs show that mutation biases are an important determinant of the prevalence for different types of resistant strains or tumors.[4][5] Thus, a knowledge of mutation bias can be used to design more evolution-resistant therapies.[4]
When mutation bias is invoked as a possible cause of some pattern of asymmetry in evolution, the alternative hypotheses may include selection, biased gene conversion, and demographic factors.
In the past, due to the technical difficulty of detecting rare mutations, most attempts to characterize the mutation spectrum were based on reporter gene systems, or based on patterns of presumptively neutral change in pseudogenes. More recently, there has been an effort to use the MA (mutation accumulation) method and high-throughput sequencing (e.g.,[6] ).
Examples of mutation biases
Transition-transversion bias
The canonical DNA nucleotides include 2 purines (A and G) and 2 pyrimidines (T and C). In the molecular evolution literature, the term transition is used for nucleotide changes within a chemical class, and transversion for changes from one chemical class to the other. Each nucleotide is subject to one transition (e.g., T to C) and 2 transversions (e.g., T to A or T to G).
Because a site (or a sequence) is subject to twice as many transversions as transitions, the total rate of transversions for a sequence may be higher even when the rate of transitions is higher on a per-path basis. In the molecular evolution literature, the per-path rate bias is typically denoted by κ (kappa), so that, if the rate of each transversion is u, the rate of each transition is κu. Then, the aggregate rate ratio (transitions to transversions) is R = (1 * κu) / (2 * u) = κ / 2. For instance, in yeast, κ ~ 1.2,[7] therefore the aggregate bias is R = 1.2 / 2 = 0.6, whereas in E. coli, κ ~ 4 so that R ~ 2.
In a variety of organisms, transition mutations occur several-fold more frequently than expected under uniformity.[8] The bias in animal viruses is sometimes much more extreme, e.g., 31 of 34 nucleotide mutations in a recent study in HIV were transitions.[9] As noted above, the bias toward transitions is weak in yeast, and appear to be absent in the grasshopper Podisma pedestris.[10]
Definition
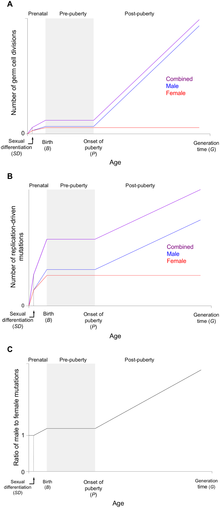
Male mutation bias is also called "Male-Driven Evolution". The rate of male germline mutations is generally higher than in females.[11] The phenomenon of Male mutation bias have been observed in many species.[12]
Origin
In 1935, the British-Indian scientist J.B.S. Haldane found that in hemophilia, the blood clotting disorder originated on the X chromosomes is due to fathers' germline mutation.[13] Then he proposed the hypothesis that the male germline contributes inordinately more mutations to succeeding generations than that in the female germline mutation.[14]
Evidence
In 1987, Takashi Miyata at al. designed an approach to test Haldane’s hypothesis.[15] If α is the ratio of the male mutation rate to the female mutation rate, Y and X are denoted as Y and X-linked sequence mutation rate, he include that the ratio of Y-linked sequence mutation rate to X-linked sequence mutation rate is:

The mean Y/X ratio is 2.25 in higher primates.[16] By using the equation, we could estimate the ratio of male to female mutation rates α ≈ 6. In some organisms with a shorter generation time than humans, the mutation rate in males is also larger than those in females. Because their cell divisions in males are usually not that large. The ratio of the number of germ cell divisions from one generation to the next in males to females is less than that in human.[17][18][19]
There are also other hypotheses that want to explain the male mutation bias. They think it may be caused by the mutation rate in the Y-linked sequence higher than the X-linked sequence mutation rate. The male germline genome is heavily methylated and more inclined to mutate than females. X chromosomes experience more purifying selection mutations on hemizygous chromosomes.[20] To test this hypothesis, people use birds to study their mutation rate.[21][22] Contrary to humans, bird males are homogametes (WW), and females are heterogametes (WZ). They found that the bird male-to-female ratio in mutation rates ranges from 4 to 7.[23] It also proved that the mutation bias is mostly resulted from more male germline mutation than the female.
Explanation
A mutation is a heritable variation in the genetic information of a short region of DNA sequences. Mutations can be categorized into replication-dependent mutations and replication-independent mutations. Therefore, there are two kinds of mutation mechanisms to explain the phenomenon of male mutation bias.
Replication-dependent mechanism
The number of germ cell divisions in females are constant and are much less than that in males. In females, most primary oocytes are formed at birth. The number of cell divisions occurred in the production of a mature ovum is constant. In males, more cell divisions are required during the process of spermatogenesis. Not only that, the cycle of spermatogenesis is never-ending. Spermatogonia will continue to divide throughout the whole productive life of the male. The number of male germline cell divisions at production is not only higher than female germline cell divisions but also mounting as the age of the male increases.[24]
One might expect the male mutation rate would be similar to the rate of male germline cell divisions. But only few species conform to the estimation of the male mutation rate.[19] Even in these species, the ratio of male-to-female mutation rate is lower than the ratio of male-to-female in the number of germline cell divisions.[25]
Replication-independent mechanism
The skew estimates of the male-to-female mutation rate ratio introduce the other important mechanism that highly influences male mutation bias. Mutations at CpG sites result in a C-to-T transition.[26] These C-to-T nucleotide substitutions occur 10-50 times faster than that at rest sites in DNA sequences, especially likely appeared in the male and female germlines.[27] The CpG mutation barely expresses any sex biases because of the independence of replication, and effectively lower the ratio of male-to-female mutation rate.[28] Besides, neighbor-dependent mutations can also cause biases in mutation rate ,and may have no relevance to DNA replication. For example, if mutations originated by the effect of mutagens show weak male mutation bias, such as exposure to the UV light.[29]
In summary, male mutation bias primarily is due to replication-dependent mutations occurred in the male germline more than in the female germline, but replication-independent mutations also contribute to alleviating the difference.
GC-AT bias
A GC-AT bias is a bias with a net effect on GC content. For instance, if G and C sites are simply more mutable than A and T sites, other things being equal, this would result in a net downward pressure on GC content.
Mutation-accumulation studies in yeast have indicated a bias toward AT of roughly 2-fold.[7]
A common idea in the literature of molecular evolution is that codon usage and genome composition reflect the effects of mutation bias, e.g., codon usage has been treated with a mutation-selection-drift model combining mutation biases, selection for translationally preferred codons, and drift.[30] To the extent that mutation bias prevails under this model, mutation bias toward GC is responsible for genomes with high GC content, and likewise the opposite bias is responsible for genomes with low GC content.
Starting in the 1990s, it became clear that GC-biased gene conversion was a major factor—previously unanticipated—in affecting GC content in diploid organisms such as mammals.[31]
Similarly, although it may be the case that bacterial genome composition strongly reflects GC and AT biases, the proposed mutational biases have not been demonstrated to exist. Indeed, Hershberg and Petrov [2] suggest that mutation in most bacterial genomes is biased toward AT, even when the genome is not AT-rich. Thus, the importance of GC-AT biases in accounting for compositional effects has not been established and is an area of ongoing research.
Other mutation biases
- Phenotypic variation may show biases (see Developmental bias)
- STR loci may exhibit biases to expand or contract [3]
- In mammals and birds, CpG sites are mutation hotspots
- Flanking nucleotides affect mutation rate in mammals [32]
- Transcription enhances mutation in a strand-specific manner [33]
Related concepts
The concept of mutation bias, as defined above, does not imply foresight, design, or even a specially evolved tendency, e.g., the bias may emerge simply as a side-effect of DNA repair processes. Currently there is no established terminology for mutation-generating systems that tend to produce useful mutations. The term "directed mutation" or adaptive mutation is sometimes used with the implication of a process of mutation that senses and responds to conditions directly. When the sense is simply that the mutation system is tuned to enhance the production of helpful mutations under certain conditions, the terminology of "mutation strategies" [34] or "natural genetic engineering" [35] has been suggested, but these terms are not widely used. Various mechanisms of mutation in pathogenic microbes, e.g., mechanisms for phase variation and antigenic variation, appear to have evolved so as to enhance lineage survival, and these mechanisms are routinely described as strategies or adaptations in the microbial genetics literature (e.g.,[36]).
References
- D. Ostrow, N. Phillips, A. Avalos, D. Blanton, A. Boggs, T. Keller, L. Levy, J. Rosenbloom and C. F. Baer (2007). "Mutational bias for body size in rhabditid nematodes". Genetics. 176 (3): 1653–61. doi:10.1534/genetics.107.074666. PMC 1931521. PMID 17483403.CS1 maint: multiple names: authors list (link)
- R. Hershberg and D. A. Petrov (2010). "Evidence that mutation is universally biased towards AT in bacteria". PLoS Genet. 6 (9): e1001115. doi:10.1371/journal.pgen.1001115. PMC 2936535. PMID 20838599.
- H. Ellegren (2000). "Microsatellite mutations in the germline: implications for evolutionary inference". Trends Genet. 16 (12): 551–8. doi:10.1016/S0168-9525(00)02139-9. PMID 11102705.
- C. Liu, S. Leighow, H. Inam, B. Zhao and J. R. Pritchard (2019). "Exploiting the 'survival of the likeliest' to enable evolution-guided drug design". bioRxiv: 557645. doi:10.1101/557645.CS1 maint: multiple names: authors list (link)
- V. L. Cannataro, S. G. Gaffney and J. P. Townsend (2018). "Effect Sizes of Somatic Mutations in Cancer". J Natl Cancer Inst. 110 (11): 1171–1177. doi:10.1093/jnci/djy168. PMC 6235682. PMID 30365005.
- M. L. Weng, C. Becker, J. Hildebrandt, M. Neumann, M. T. Rutter, R. G. Shaw, D. Weigel and C. B. Fenster (2019). "Fine-Grained Analysis of Spontaneous Mutation Spectrum and Frequency in Arabidopsis thaliana". Genetics. 211 (2): 703–714. doi:10.1534/genetics.118.301721. PMC 6366913. PMID 30514707.CS1 maint: multiple names: authors list (link)
- M. Lynch, W. Sung, K. Morris, N. Coffey, C. R. Landry, E. B. Dopman, W. J. Dickinson, K. Okamoto, S. Kulkarni, D. L. Hartl and W. K. Thomas (2008). "A genome-wide view of the spectrum of spontaneous mutations in yeast". Proc Natl Acad Sci U S A. 105 (27): 9272–7. doi:10.1073/pnas.0803466105. PMC 2453693. PMID 18583475.CS1 maint: multiple names: authors list (link)
- A. Stoltzfus and R. W. Norris (2016). "On the Causes of Evolutionary Transition:Transversion Bias". Mol Biol Evol. 33 (3): 595–602. doi:10.1093/molbev/msv274. PMID 26609078.
- P. Yap, D. W.-S. Koh, C. T.-T. Su, K.-F. Chan and S. K.-E. Gan (2019). "Predicting mutations in HIV-1 Gag: Insights from in silico and an in vitro BSL2 platform on thermostability and allosteric effects". bioRxiv: 679852. doi:10.1101/679852.CS1 maint: multiple names: authors list (link)
- I. Keller, D. Bensasson and R. A. Nichols (2007). "Transition-transversion bias is not universal: a counter example from grasshopper pseudogenes". PLoS Genet. 3 (2): e22. doi:10.1371/journal.pgen.0030022. PMC 1790724. PMID 17274688.
- Li, W (2002-12-01). "Male-driven evolution". Current Opinion in Genetics & Development. 12 (6): 650–656. doi:10.1016/s0959-437x(02)00354-4. ISSN 0959-437X. PMID 12433577.
- Sloan, Daniel Benjamin. Mutation Rate Variation and Organelle Genome Evolution in the Angiosperm Genus Silene (Thesis). University of Virginia. doi:10.18130/v3rp1d.
- Haldane, J. B. S. (October 1935). "The rate of spontaneous mutation of a human gene". Journal of Genetics. 31 (3): 317–326. doi:10.1007/bf02982403. ISSN 0022-1333.
- HALDANE, J. B. S. (January 1946). "The Mutation Rate of the Gene for Haemophilia, and ITS Segregation Ratios in Males and Females". Annals of Eugenics. 13 (1): 262–271. doi:10.1111/j.1469-1809.1946.tb02367.x. ISSN 2050-1420.
- Miyata, T.; Hayashida, H.; Kuma, K.; Mitsuyasu, K.; Yasunaga, T. (1987-01-01). "Male-driven Molecular Evolution: A Model and Nucleotide Sequence Analysis". Cold Spring Harbor Symposia on Quantitative Biology. 52: 863–867. doi:10.1101/sqb.1987.052.01.094. ISSN 0091-7451.
- Shimmin, Lawrence C.; Chang, Benny Hung-Junn; Hewett-Emmett, David; Li, Wen-Hsiung (August 1993). "Potential problems in estimating the male-to-female mutation rate ratio from DNA sequence data". Journal of Molecular Evolution. 37 (2): 160–166. Bibcode:1993JMolE..37..160S. doi:10.1007/bf02407351. ISSN 0022-2844. PMID 8411204.
- Chang, B. H.; Shimmin, L. C.; Shyue, S. K.; Hewett-Emmett, D.; Li, W. H. (1994-01-18). "Weak male-driven molecular evolution in rodents". Proceedings of the National Academy of Sciences. 91 (2): 827–831. Bibcode:1994PNAS...91..827C. doi:10.1073/pnas.91.2.827. ISSN 0027-8424. PMID 8290607.
- "The Zuckerkandl Prize". Journal of Molecular Evolution. 56 (4): 373–374. April 2003. Bibcode:2003JMolE..56..373.. doi:10.1007/s00239-002-2455-5. ISSN 0022-2844.
- Sayres, Melissa A. Wilson; Venditti, Chris; Pagel, Mark; Makova, Kateryna D. (October 2011). "Do Variations in Substitution Rates and Male Mutation Bias Correlate with Life-History Traits? A Study of 32 Mammalian Genomes". Evolution. 65 (10): 2800–2815. doi:10.1111/j.1558-5646.2011.01337.x. PMID 21967423.
- McVean, Gilean T.; Hurst, Laurence D. (March 1997). "Evidence for a selectively favourable reduction in the mutation rate of the X chromosome". Nature. 386 (6623): 388–392. Bibcode:1997Natur.386..388M. doi:10.1038/386388a0. ISSN 0028-0836.
- Ellegren, Hans; Fridolfsson, Anna-Karin (October 1997). "Male–driven evolution of DNA sequences in birds". Nature Genetics. 17 (2): 182–184. doi:10.1038/ng1097-182. ISSN 1061-4036.
- Axelsson, Erik; Smith, Nick G.C.; Sundström, Hannah; Berlin, Sofia; Ellegren, Hans (August 2004). "Male-Biased Mutation Rate and Divergence in Autosomal, Z-Linked and W-Linked Introns of Chicken and Turkey". Molecular Biology and Evolution. 21 (8): 1538–1547. doi:10.1093/molbev/msh157. ISSN 1537-1719.
- Smeds, Linnéa; Qvarnström, Anna; Ellegren, Hans (2016-07-13). "Direct estimate of the rate of germline mutation in a bird". Genome Research. 26 (9): 1211–1218. doi:10.1101/gr.204669.116. ISSN 1088-9051.
- Elsas, Louis J. (December 1981). "Human Genetics Human Genetics: Problems and Approaches Vogel Motulsky". BioScience. 31 (11): 847. doi:10.2307/1308691. ISSN 0006-3568. JSTOR 1308691.
- Wilson Sayres, Melissa A.; Makova, Kateryna D. (2011-10-18). "Genome analyses substantiate male mutation bias in many species". BioEssays. 33 (12): 938–945. doi:10.1002/bies.201100091. ISSN 0265-9247. PMC 4600401. PMID 22006834.
- Ehrlich, M; Wang, R. (1981-06-19). "5-Methylcytosine in eukaryotic DNA". Science. 212 (4501): 1350–1357. Bibcode:1981Sci...212.1350E. doi:10.1126/science.6262918. ISSN 0036-8075.
- Walser, J.-C.; Furano, A. V. (2010-05-24). "The mutational spectrum of non-CpG DNA varies with CpG content". Genome Research. 20 (7): 875–882. doi:10.1101/gr.103283.109. ISSN 1088-9051.
- Drozdov, A. L. (March 2006). "Academician Vladimir Leonidovich Kas'yanov". Russian Journal of Marine Biology. 32 (1): 71–73. doi:10.1134/s1063074006010111. ISSN 1063-0740.
- Arndt, P. F.; Hwa, T. (2005-03-15). "Identification and measurement of neighbor-dependent nucleotide substitution processes". Bioinformatics. 21 (10): 2322–2328. arXiv:q-bio/0501018. Bibcode:2005q.bio.....1018A. doi:10.1093/bioinformatics/bti376. ISSN 1367-4803.
- M. Bulmer (1991). "The selection-mutation-drift theory of synonymous codon usage". Genetics. 129 (3): 897–907. PMC 1204756. PMID 1752426.
- L. Duret and N. Galtier (2009). "Biased gene conversion and the evolution of mammalian genomic landscapes". Annu Rev Genom Hum Genet. 10: 285–311. doi:10.1146/annurev-genom-082908-150001. PMID 19630562.
- D. G. Hwang and P. Green (2004). "Bayesian Markov chain Monte Carlo sequence analysis reveals varying neutral substitution patterns in mammalian evolution". Proc Natl Acad Sci U S A. 101 (39): 13994–4001. Bibcode:2004PNAS..10113994H. doi:10.1073/pnas.0404142101. PMC 521089. PMID 15292512.
- N. Kim and S. Jinks-Robertson (2012). "Transcription as a source of genome instability". Nat Rev Genet. 13 (3): 204–14. doi:10.1038/nrg3152. PMC 3376450. PMID 22330764.
- L. H. Caporale (2003). Darwin in the Genome: Molecular strategies in biological evolution. McGraw-Hill.
- J. Shapiro (2011). Evolution: A View From the 21st Century. FT Press, New York.
- J. Foley (2015). "Mini-review: Strategies for Variation and Evolution of Bacterial Antigens". Comput Struct Biotechnol J. 13: 407–16. doi:10.1016/j.csbj.2015.07.002. PMID 26288700.
