Notch proteins
Notch proteins are a family of type-1 transmembrane proteins that form a core component of the Notch signaling pathway, which is highly conserved in metazoans. The Notch extracellular domain (NECD) mediates interactions with DSL family ligands, allowing it to participate in juxtacrine signaling.The Notch intracellular domain (NICD) acts as a transcriptional activator when in complex with CSL family transcription factors. Members of this Type 1 transmembrane protein family share several core structures, including an extracellular domain consisting of multiple epidermal growth factor (EGF)-like repeats and an intracellular domain transcriptional activation domain (TAD). Notch family members operate in a variety of different tissues and play a role in a variety of developmental processes by controlling cell fate decisions. Much of what is known about Notch function comes from studies done in Caenorhabditis elegans (C.elegans) and Drosophila melanogaster. Human homologs have also been identified, but details of Notch function and interactions with its ligands are not well known in this context.
| Notch (LNR) domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| Symbol | Notch | ||||||||
| Pfam | PF00066 | ||||||||
| InterPro | IPR000800 | ||||||||
| SMART | SM00004 | ||||||||
| PROSITE | PS50258 | ||||||||
| OPM superfamily | 462 | ||||||||
| OPM protein | 5kzo | ||||||||
| Membranome | 19 | ||||||||
| |||||||||
Discovery
Notch was discovered in a mutant Drosophila in March 1913 in the lab of Thomas Hunt Morgan.[2] This mutant emerged after several generations of crossing out and back-crossing beaded winged flies with wild type flies and was first characterized by John S. Dexter.[3] The most frequently observed phenotype in Notch mutant flies is the appearance of a concave serration at the most distal end of the wings, for which the gene is named, accompanied by the absence of marginal bristles.[4][5] This mutant was found to be a sex-linked dominant on the X chromosome that could only be observed in heterozygous females as it was lethal in males and homozygous females.[2] The first Notch allele was established in 1917 by C.W. Metz and C.B. Bridges.[6] In the late 1930s, studies of fly embryogenesis done by Donald F. Poulson provided the first indication of Notch's role in development.[7] Notch-8 mutant males exhibited a lack of the inner germ layers, the endoderm and mesoderm, that resulted in failure to undergo later morphogenesis embryonic lethality. Later studies in early Drosophila neurogenesis provided some of the first indications of Notch's roll in cell-cell signaling, as the nervous system in Notch mutants was developed by sacrificing hypodermal cells.[8]
Starting in the 1980s researchers began to gain further insights into Notch function through genetic and molecular experiments. Genetic screens conducted in Drosophila led to the identification of several proteins that play a central role in Notch signaling, including Enhancer of split,[8] Master mind, Delta,[9] Suppressor of Hairless (CSL),[10] and Serrate.[11] At the same time, the Notch gene was successfully sequenced[12][13] and cloned,[14][15] providing insights into the molecular architecture of Notch proteins and led to identification of Notch homologs in Caenorhabditis elegans (C. elegans)[16][17][18] and eventually in mammals.
In the early 1990s Notch was increasingly implicated as the receptor of a previously unknown intercellular signal pathway[19][20] in which the NICD is transported to the nucleus where it acts as a transcription factor to directly regulate target genes.[21][22][23] The release of the NICD was found to be as a result of proteolytic cleavage of the transmembrane protein through the actions of the γ-secretase complex catalytic subunit Presenilin. This was a significant interaction as Presenilin is implicated in the development of Alzheimer's disease.[24] This and further research into the mechanism of Notch signaing led to research that would further connect Notch to a wide range of human diseases.
Structure
Drosophila contain a single Notch protein, C. elegans contain two redundant notch paralogs, Lin-12[25] and GLP-1,[18][26] and humans have four Notch variants, Notch 1-4. Although variations exist between homologs, there are a set of highly conserved structures found in all Notch family proteins. The protein can broadly be split into the Notch extracellular domain (NECD) and Notch intracellular domain (NICD) joined together by a single-pass transmembrane domain (TM).
The NECD contains 36 EGF repeats in Drosophila,[13] 28-36 in humans, and 13 and 10 in C. elegans Lin-12 and GLP-1 respectively.[27] These repeats are heavily modified through O-glycoslyation[28] and the addition of specific O-linked glycans has been shown to be necessary for proper function. The EGF repeats are followed by three cysteine-rich Lin-12/Notch Repeats (LNR) and a heterodimerization (HD) domain. Together the LNR and HD compose the negative regulatory region adjacent to the cell membrane and help prevent signaling in the absence of ligand binding.
The NICD acts as a transcription factor that is released after ligand binding triggers its cleavage. It contains a nuclear localization sequence (NLS) that mediates its translocation to the nucleus. where it forms a transcriptional complex along with several other transcription factors. Once in the nucleus, several akyrin repeats (ANK) and the RAM domain interactions between the NICD and CSL proteins to form a transcriptional activation complex.[29] In humans, an additional PEST domain plays a role in NICD degradation.[30]
Function
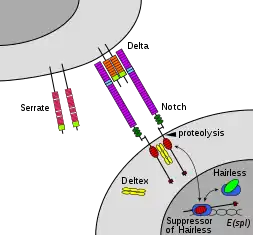
Notch family members play a role in a variety of developmental processes by controlling cell fate decisions. The Notch signaling network is an evolutionarily conserved intercellular signaling pathway that regulates interactions between physically adjacent cells. In Drosophila, notch interaction with its cell-bound ligands (delta, serrate) establishes an intercellular signaling pathway that plays a key role in development. This protein functions as a receptor for membrane bound ligands, and may play multiple roles during development.[31] A deficiency can be associated with bicuspid aortic valve.[32]
There is evidence that activated Notch 1 and Notch 3 promote differentiation of progenitor cells into astroglia.[33] Notch 1, then activated before birth, induces radial glia differentiation,[34] but postnatally induces the differentiation into astrocytes.[35] One study shows that Notch-1 cascade is activated by Reelin in an unidentified way.[36] Reelin and Notch1 cooperate in the development of the dentate gyrus, according to another.[37]
Ligand interactions
| Jagged/Serrate protein | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | DSL | ||||||||
| Pfam | PF01414 | ||||||||
| InterPro | IPR026219 | ||||||||
| Membranome | 76 | ||||||||
| |||||||||
Notch signaling is triggered via direct cell-to-cell contact, mediated by interactions between the Notch receptor protein in the signal receiving cell and a ligand in an adjacent signal transmitting cell. These type 1 single pass transmembrane proteins fall into the Delta/Serrate/Lag-2 (DSL) family of proteins which is named after the three canonical Notch ligands.[19] Delta and Serrate are found in Drosophila while Lag-2 is found in C. elegans. Humans contain 3 Delta homologs, Delta-like 1, 3, and 4, as well as two Serrate homolgs, Jagged 1 and 2. Notch proteins consist of a relatively short intracellular domain and a large extracellular domain with one or more EGF motifs and a N-terminal DSL-binding motif. EGF repeats 11-12 on the Notch extracellular domain have been shown to be necessary and sufficient for trans signaling interactions between Notch and its ligands.[38] Additionally, EGF repeats 24-29 have been implicated in inhibition of cis interactions between Notch and ligands co-expressed in the same cell.[39]
Proteolysis
In order for a signaling event to occur, the Notch protein must be cleaved at several sites. In humans, Notch is first cleaved in the NRR domain by Furin while being processed in the trans-Golgi network before being presented on the cell surface as a heterodimer.[40][41] Drosophila Notch does not require this cleavage for signaling to occur,[42] and there is some evidence that suggests that LIN-12 and GLP-1 are cleaved at this site in C. elegans.
Release of the NICD is achieved after an additional two cleavage events to Notch. Binding of Notch to a DSL ligand results in a conformational change that exposes a cleavage site in the NECD. Enzymatic proteolysis at this site is carried out by a A Disintegrin and Metalloprotease domain (ADAM) family protease. This protein is called Kuzbanian in Drosopihla,[43][44] sup-17 in C. elegans,[45] and ADAM10 in humans.[46][47] After proteolytic cleavage, the released NECD is endocytosed into the signal transmitting cell, leaving behind only a small extracellular portion of Notch. This truncated Notch protein can then be recognized by a γ-secretase that cleaves the third site found in the TM domain.[48]
Human homologs
Notch-1
Notch-2
Notch-2 (Neurogenic locus notch homolog protein 2) is a protein that in humans is encoded by the NOTCH2 gene.[49]
NOTCH2 is associated with Alagille syndrome[50] and Hajdu–Cheney syndrome.[51]
Notch-3
Notch-4
See also
Notes
- Vardar D, North CL, Sanchez-Irizarry C, Aster JC, Blacklow SC (June 2003). "Nuclear magnetic resonance structure of a prototype Lin12-Notch repeat module from human Notch1". Biochemistry. 42 (23): 7061–7. doi:10.1021/bi034156y. PMID 12795601.
- Morgan TH, Bridges CB (1916). Sex-linked inheritance in Drosophila. NCSU Libraries. Washington, Carnegie Institution of Washington.
- Dexter JS (December 1914). "The Analysis of a Case of Continuous Variation in Drosophila by a Study of Its Linkage Relations". The American Naturalist. 48 (576): 712–758. doi:10.1086/279446.
- Mohr OL (May 1919). "Character Changes Caused by Mutation of an Entire Region of a Chromosome in Drosophila". Genetics. 4 (3): 275–82. doi:10.1093/genetics/4.3.275. PMC 1200460. PMID 17245926.
- Lindsley DL, Zimm GG (2012-12-02). The Genome of Drosophila Melanogaster. Academic Press. ISBN 9780323139847.
- Metz CW, Bridges CB (December 1917). "Incompatibility of Mutant Races in Drosophila". Proceedings of the National Academy of Sciences of the United States of America. 3 (12): 673–8. Bibcode:1917PNAS....3..673M. doi:10.1073/pnas.3.12.673. PMC 1091355. PMID 16586764.
- Poulson DF (March 1937). "Chromosomal Deficiencies and the Embryonic Development of Drosophila Melanogaster". Proceedings of the National Academy of Sciences of the United States of America. 23 (3): 133–7. Bibcode:1937PNAS...23..133P. doi:10.1073/pnas.23.3.133. PMC 1076884. PMID 16588136.
- Lehmann R, Jiménez F, Dietrich U, Campos-Ortega JA (March 1983). "On the phenotype and development of mutants of early neurogenesis inDrosophila melanogaster". Wilhelm Roux's Archives of Developmental Biology. 192 (2): 62–74. doi:10.1007/BF00848482. PMID 28305500. S2CID 25602190.
- Lehmann R, Dietrich U, Jiménez F, Campos-Ortega JA (July 1981). "Mutations of early neurogenesis inDrosophila". Wilhelm Roux's Archives of Developmental Biology. 190 (4): 226–229. doi:10.1007/BF00848307. PMID 28305572. S2CID 21814447.
- Fortini ME, Artavanis-Tsakonas S (October 1994). "The suppressor of hairless protein participates in notch receptor signaling". Cell. 79 (2): 273–82. doi:10.1016/0092-8674(94)90196-1. PMID 7954795. S2CID 40771329.
- Fleming RJ, Scottgale TN, Diederich RJ, Artavanis-Tsakonas S (December 1990). "The gene Serrate encodes a putative EGF-like transmembrane protein essential for proper ectodermal development in Drosophila melanogaster". Genes & Development. 4 (12A): 2188–201. doi:10.1101/gad.4.12a.2188. PMID 2125287.
- Kidd S, Kelley MR, Young MW (September 1986). "Sequence of the notch locus of Drosophila melanogaster: relationship of the encoded protein to mammalian clotting and growth factors". Molecular and Cellular Biology. 6 (9): 3094–108. doi:10.1128/mcb.6.9.3094. PMC 367044. PMID 3097517.
- Wharton KA, Johansen KM, Xu T, Artavanis-Tsakonas S (December 1985). "Nucleotide sequence from the neurogenic locus notch implies a gene product that shares homology with proteins containing EGF-like repeats". Cell. 43 (3 Pt 2): 567–81. doi:10.1016/0092-8674(85)90229-6. PMID 3935325.
- Kidd S, Lockett TJ, Young MW (September 1983). "The Notch locus of Drosophila melanogaster". Cell. 34 (2): 421–33. doi:10.1016/0092-8674(83)90376-8. PMID 6193889. S2CID 36425372.
- Artavanis-Tsakonas S, Muskavitch MA, Yedvobnick B (April 1983). "Molecular cloning of Notch, a locus affecting neurogenesis in Drosophila melanogaster". Proceedings of the National Academy of Sciences of the United States of America. 80 (7): 1977–81. Bibcode:1983PNAS...80.1977A. doi:10.1073/pnas.80.7.1977. PMC 393735. PMID 6403942.
- Greenwald I (February 1987). "The lin-12 locus of Caenorhabditis elegans". BioEssays. 6 (2): 70–3. doi:10.1002/bies.950060207. PMID 3551950. S2CID 19253030.
- Priess JR, Schnabel H, Schnabel R (November 1987). "The glp-1 locus and cellular interactions in early C. elegans embryos". Cell. 51 (4): 601–11. doi:10.1016/0092-8674(87)90129-2. PMID 3677169. S2CID 6282210.
- Austin J, Kimble J (November 1987). "glp-1 is required in the germ line for regulation of the decision between mitosis and meiosis in C. elegans". Cell. 51 (4): 589–99. doi:10.1016/0092-8674(87)90128-0. PMID 3677168. S2CID 31484517.
- Artavanis-Tsakonas S, Matsuno K, Fortini ME (April 1995). "Notch signaling". Science. 268 (5208): 225–32. Bibcode:1995Sci...268..225A. doi:10.1126/science.7716513. PMID 7716513.
- Greenwald I, Rubin GM (January 1992). "Making a difference: the role of cell-cell interactions in establishing separate identities for equivalent cells". Cell. 68 (2): 271–81. doi:10.1016/0092-8674(92)90470-w. PMID 1365402. S2CID 11901508.
- Schroeter EH, Kisslinger JA, Kopan R (May 1998). "Notch-1 signalling requires ligand-induced proteolytic release of intracellular domain". Nature. 393 (6683): 382–6. Bibcode:1998Natur.393..382S. doi:10.1038/30756. PMID 9620803. S2CID 4431882.
- Kopan R, Nye JS, Weintraub H (September 1994). "The intracellular domain of mouse Notch: a constitutively activated repressor of myogenesis directed at the basic helix-loop-helix region of MyoD". Development. 120 (9): 2385–96. PMID 7956819.
- Struhl G, Fitzgerald K, Greenwald I (July 1993). "Intrinsic activity of the Lin-12 and Notch intracellular domains in vivo". Cell. 74 (2): 331–45. doi:10.1016/0092-8674(93)90424-o. PMID 8343960. S2CID 27966283.
- Sherrington R, Rogaev EI, Liang Y, Rogaeva EA, Levesque G, Ikeda M, et al. (June 1995). "Cloning of a gene bearing missense mutations in early-onset familial Alzheimer's disease". Nature. 375 (6534): 754–60. Bibcode:1995Natur.375..754S. doi:10.1038/375754a0. PMID 7596406. S2CID 4308372.
- Greenwald IS, Sternberg PW, Horvitz HR (September 1983). "The lin-12 locus specifies cell fates in Caenorhabditis elegans". Cell. 34 (2): 435–44. doi:10.1016/0092-8674(83)90377-x. PMID 6616618. S2CID 40668388.
- Austin J, Kimble J (August 1989). "Transcript analysis of glp-1 and lin-12, homologous genes required for cell interactions during development of C. elegans". Cell. 58 (3): 565–71. doi:10.1016/0092-8674(89)90437-6. PMID 2758467. S2CID 8514017.
- Greenwald I (December 1985). "lin-12, a nematode homeotic gene, is homologous to a set of mammalian proteins that includes epidermal growth factor". Cell. 43 (3 Pt 2): 583–90. doi:10.1016/0092-8674(85)90230-2. PMID 3000611.
- Shao L, Luo Y, Moloney DJ, Haltiwanger R (November 2002). "O-glycosylation of EGF repeats: identification and initial characterization of a UDP-glucose: protein O-glucosyltransferase". Glycobiology. 12 (11): 763–70. doi:10.1093/glycob/cwf085. PMID 12460944.
- Tamura K, Taniguchi Y, Minoguchi S, Sakai T, Tun T, Furukawa T, Honjo T (December 1995). "Physical interaction between a novel domain of the receptor Notch and the transcription factor RBP-J kappa/Su(H)". Current Biology. 5 (12): 1416–23. doi:10.1016/S0960-9822(95)00279-X. hdl:2433/202204. PMID 8749394. S2CID 18442572.
- Weng AP, Ferrando AA, Lee W, Morris JP, Silverman LB, Sanchez-Irizarry C, et al. (October 2004). "Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia". Science. 306 (5694): 269–71. Bibcode:2004Sci...306..269W. doi:10.1126/science.1102160. PMID 15472075. S2CID 24049536.
- "Entrez Gene: NOTCH1 Notch homolog 1, translocation-associated (Drosophila)".
- McKellar SH, Tester DJ, Yagubyan M, Majumdar R, Ackerman MJ, Sundt TM (August 2007). "Novel NOTCH1 mutations in patients with bicuspid aortic valve disease and thoracic aortic aneurysms". The Journal of Thoracic and Cardiovascular Surgery. 134 (2): 290–6. doi:10.1016/j.jtcvs.2007.02.041. PMID 17662764.
- Tanigaki K, Nogaki F, Takahashi J, Tashiro K, Kurooka H, Honjo T (January 2001). "Notch1 and Notch3 instructively restrict bFGF-responsive multipotent neural progenitor cells to an astroglial fate". Neuron. 29 (1): 45–55. doi:10.1016/S0896-6273(01)00179-9. hdl:2433/150564. PMID 11182080. S2CID 17047028.
- Gaiano N, Nye JS, Fishell G (May 2000). "Radial glial identity is promoted by Notch1 signaling in the murine forebrain". Neuron. 26 (2): 395–404. doi:10.1016/S0896-6273(00)81172-1. PMID 10839358. S2CID 15861936.
- Chambers CB, Peng Y, Nguyen H, Gaiano N, Fishell G, Nye JS (March 2001). "Spatiotemporal selectivity of response to Notch1 signals in mammalian forebrain precursors". Development. 128 (5): 689–702. PMID 11171394.
- Keilani S, Sugaya K (July 2008). "Reelin induces a radial glial phenotype in human neural progenitor cells by activation of Notch-1". BMC Developmental Biology. 8 (1): 69. doi:10.1186/1471-213X-8-69. PMC 2447831. PMID 18593473.
- Sibbe M, Förster E, Basak O, Taylor V, Frotscher M (July 2009). "Reelin and Notch1 cooperate in the development of the dentate gyrus". The Journal of Neuroscience. 29 (26): 8578–85. doi:10.1523/JNEUROSCI.0958-09.2009. PMC 6665659. PMID 19571148.
- Rebay I, Fleming RJ, Fehon RG, Cherbas L, Cherbas P, Artavanis-Tsakonas S (November 1991). "Specific EGF repeats of Notch mediate interactions with Delta and Serrate: implications for Notch as a multifunctional receptor". Cell. 67 (4): 687–99. doi:10.1016/0092-8674(91)90064-6. PMID 1657403. S2CID 12643727.
- de Celis JF, Bray SJ (March 2000). "The Abruptex domain of Notch regulates negative interactions between Notch, its ligands and Fringe". Development. 127 (6): 1291–302. PMID 10683181.
- Blaumueller CM, Qi H, Zagouras P, Artavanis-Tsakonas S (July 1997). "Intracellular cleavage of Notch leads to a heterodimeric receptor on the plasma membrane". Cell. 90 (2): 281–91. doi:10.1016/s0092-8674(00)80336-0. PMID 9244302. S2CID 16544864.
- Logeat F, Bessia C, Brou C, LeBail O, Jarriault S, Seidah NG, Israël A (July 1998). "The Notch1 receptor is cleaved constitutively by a furin-like convertase". Proceedings of the National Academy of Sciences of the United States of America. 95 (14): 8108–12. Bibcode:1998PNAS...95.8108L. doi:10.1073/pnas.95.14.8108. PMC 20937. PMID 9653148.
- Kidd S, Lieber T (July 2002). "Furin cleavage is not a requirement for Drosophila Notch function". Mechanisms of Development. 115 (1–2): 41–51. doi:10.1016/s0925-4773(02)00120-x. PMID 12049766. S2CID 10249987.
- Rooke J, Pan D, Xu T, Rubin GM (August 1996). "KUZ, a conserved metalloprotease-disintegrin protein with two roles in Drosophila neurogenesis". Science. 273 (5279): 1227–31. Bibcode:1996Sci...273.1227R. doi:10.1126/science.273.5279.1227. PMID 8703057. S2CID 10984937.
- Pan D, Rubin GM (July 1997). "Kuzbanian controls proteolytic processing of Notch and mediates lateral inhibition during Drosophila and vertebrate neurogenesis". Cell. 90 (2): 271–80. doi:10.1016/s0092-8674(00)80335-9. PMID 9244301. S2CID 18819481.
- Wen C, Metzstein MM, Greenwald I (December 1997). "SUP-17, a Caenorhabditis elegans ADAM protein related to Drosophila KUZBANIAN, and its role in LIN-12/NOTCH signalling". Development. 124 (23): 4759–67. PMID 9428412.
- Howard L, Glynn P (1995). "Membrane-associated metalloproteinase recognized by characteristic cleavage of myelin basic protein: assay and isolation". Methods in Enzymology. 248: 388–95. doi:10.1016/0076-6879(95)48025-0. PMID 7545777.
- Lunn CA, Fan X, Dalie B, Miller K, Zavodny PJ, Narula SK, Lundell D (January 1997). "Purification of ADAM 10 from bovine spleen as a TNFalpha convertase". FEBS Letters. 400 (3): 333–5. doi:10.1016/s0014-5793(96)01410-x. PMID 9009225. S2CID 83810622.
- Struhl G, Adachi A (September 2000). "Requirements for presenilin-dependent cleavage of notch and other transmembrane proteins". Molecular Cell. 6 (3): 625–36. doi:10.1016/s1097-2765(00)00061-7. PMID 11030342.
- Larsson C, Lardelli M, White I, Lendahl U (November 1994). "The human NOTCH1, 2, and 3 genes are located at chromosome positions 9q34, 1p13-p11, and 19p13.2-p13.1 in regions of neoplasia-associated translocation". Genomics. 24 (2): 253–8. doi:10.1006/geno.1994.1613. PMID 7698746.
- Samejima H, Torii C, Kosaki R, Kurosawa K, Yoshihashi H, Muroya K, Okamoto N, Watanabe Y, Kosho T, Kubota M, Matsuda O, Goto M, Izumi K, Takahashi T, Kosaki K (2007). "Screening for Alagille syndrome mutations in the JAG1 and NOTCH2 genes using denaturing high-performance liquid chromatography". Genetic Testing. 11 (3): 216–27. doi:10.1089/gte.2006.0519. PMID 17949281.
- Simpson MA, Irving MD, Asilmaz E, Gray MJ, Dafou D, Elmslie FV, Mansour S, Holder SE, Brain CE, Burton BK, Kim KH, Pauli RM, Aftimos S, Stewart H, Kim CA, Holder-Espinasse M, Robertson SP, Drake WM, Trembath RC (March 2011). "Mutations in NOTCH2 cause Hajdu-Cheney syndrome, a disorder of severe and progressive bone loss". Nature Genetics. 43 (4): 303–5. doi:10.1038/ng.779. PMID 21378985. S2CID 205357391.
References
- Tax FE, Yeargers JJ, Thomas JH (March 1994). "Sequence of C. elegans lag-2 reveals a cell-signalling domain shared with Delta and Serrate of Drosophila". Nature. 368 (6467): 150–4. Bibcode:1994Natur.368..150T. doi:10.1038/368150a0. PMID 8139658. S2CID 4345508.
- Lindsell CE, Shawber CJ, Boulter J, Weinmaster G (March 1995). "Jagged: a mammalian ligand that activates Notch1". Cell. 80 (6): 909–17. doi:10.1016/0092-8674(95)90294-5. PMID 7697721. S2CID 11720367.
- Kelley MR, Kidd S, Deutsch WA, Young MW (November 1987). "Mutations altering the structure of epidermal growth factor-like coding sequences at the Drosophila Notch locus". Cell. 51 (4): 539–48. doi:10.1016/0092-8674(87)90123-1. PMID 3119223. S2CID 34338340.