Signaling lymphocytic activation molecule
Signaling lymphocytic activation molecule (SLAM) is a family of genes. Homophilic binding between SLAMs is involved in cell-to-cell adhesion during antigen presentation. [1][2]
| Signaling lymphocytic activation molecule | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | SLAM | ||||||||
| Pfam | PF06214 | ||||||||
| InterPro | IPR010407 | ||||||||
| Membranome | 164 | ||||||||
| |||||||||
Signaling lymphocytic activation molecules are a CD2-related surface receptor expressed by activated phagocytes, T helper cells, and platelets.[3] SLAMs have a variety of functions, including enhancing T cellular proliferation by stimulating IL-4 and IFN-gamma production.[4] SLAM family (SLAMF) receptors can interact directly with microbes, which can cause phagocytic cells to migrate to the area.[5] SLAMF activation can trigger SLAM-associated protein (SAP) activation and a defective SAP can cause X-linked lymphoproliferative syndrome (XLP).[4]
Family Members
Members of the family include:
Location and Function
SLAMFs are CD2-related surface receptors expressed by activated B and T cells, natural killer (NK) cells, dendritic cells, macrophages, eosinophils, neutrophils, and platelets, although different SLAMF receptors have varying expression patterns.[4] SLAMF receptors are cell surface transmembrane molecules that can interact directly with microbes, which can cause phagocytic cells to migrate to the area.[11] SLAMF1 and SLAMF6 are known to directly interact with outer membrane porins on gram negative bacteria.[3] SLAMF1 is a known receptor for the measles virus and also serves as an opsonin for phagocytic cells, enhancing phagocytosis by localizing to phagosomes and inducing a signaling cascade resulting in enhanced fusion of phagosomes and lysosomes.[3][5] SLAMF2 is known bind gram negative bacteria and is internalized after binding, promoting phagocytosis.[3] SLAMFs are also involved in immune cell communication; SLAMFs are co-stimulatory molecules for both T-cells and NK cells. SLAMs enhance T helper cell proliferation by increasing IFN-gamma and IL-4 production. [11]
Structure
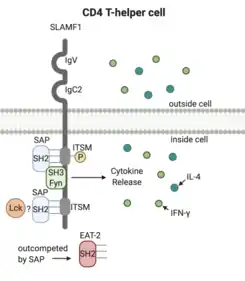
All members of the SLAMF family are classified as type I glycoproteins and share an amino-terminal IgV variable domain and a membrane-adjacent IgC2 constant domain, along with immunoreceptor tyrosine-based switch motifs (ITSMs).[11][4] The IgV and IgC2 domains are located on the extracellular portion of the receptor, while the ITSMs are used for signaling within the cell. SLAMFs can undergo alternative splicing, which can generate different isoforms of the SLAMF molecules that have different numbers of ITSMs and tyrosines, potentially with different functions.[4] Proteins with SH2 domains are able to bind these ITSMs to initiate signaling cascades within the cell. SLAMF2 and SLAMF4 interact with one another, but all other SLAMF receptors are self-ligands, meaning that they interact with the corresponding receptor on other cells in a homophilic way.[3]
Uses in Immunotherapy
SLAMFs are potential targets for immunotherapy. For example, elotuzumab is an anti-SLAMF7 humanized monoclonal antibody used to treat multiple myeloma. SLAMF7 is a self-ligand over-expressed in plasma cells of multiple myeloma patients. Elotuzumab stimulates NK cells to release granzyme through blocking SLAMF7, through triggering antibody-dependent cellular cytotoxicity (ADCC), and through NK cell activation via Ewing’s sarcoma-associated transcript 2 (EAT-2).[12] EAT-2 is known to bind to phosphorylated tyrosines on ITSMs and alter cytokine production.[5] Elotuzumab also blocks multiple myeloma cells from interacting with one another via the SLAMF7 ligand.[13]
SLAM-Associated Protein (SAP)
The X-linked SLAM-associated protein (SAP), encoded by the SH2D1A gene, consists primarily of an SH2 domain which can interact with ITSMs present on most SLAMF receptors.[3][4] Unlike most SH2 binding proteins, SAP does not require tyrosines on the ITSMs to be phosphorylated prior to binding.[4] SAP is expressed in lymphocytes (specifically NK cells and T cells, but not usually B cells), eosinophils, and platelets.[4][3] A defective SLAM associated protein (SAP) causes X-linked lymphoproliferative syndrome (XLP), a frequently lethal mononucleosis characterized by inability to respond to infection with Epstein-Barr virus (EBV), leading to a failure to clear B-cells infected with the virus, which can be fatal.[3]
References
- "SLAM FAMILY, MEMBER 1". OMIM. Retrieved 19 March 2014.
- Rosenbach T, Csatò M, Czarnetzki BM (January 1988). "Studies on the role of leukotrienes in murine allergic and irritant contact dermatitis". The British Journal of Dermatology. 118 (1): 1–6. doi:10.1111/j.1365-2133.1988.tb01743.x. PMID 2829957.
- Detre, Cynthia; Keszei, Marton; Romero, Xavier; Tsokos, George C.; Terhorst, Cox (2010-06-01). "SLAM family receptors and the SLAM-associated protein (SAP) modulate T cell functions". Seminars in Immunopathology. 32 (2): 157–171. doi:10.1007/s00281-009-0193-0. ISSN 1863-2300. PMC 2868096. PMID 20146065.
- Cannons, Jennifer L.; Tangye, Stuart G.; Schwartzberg, Pamela L. (2011-04-23). "SLAM Family Receptors and SAP Adaptors in Immunity". Annual Review of Immunology. 29 (1): 665–705. doi:10.1146/annurev-immunol-030409-101302. ISSN 0732-0582. PMID 21219180.
- van Driel BJ, Liao G, Engel P, Terhorst C (2016-01-20). "Responses to Microbial Challenges by SLAMF Receptors". Frontiers in Immunology. 7: 4. doi:10.3389/fimmu.2016.00004. PMC 4718992. PMID 26834746.
- Sintes J, Engel P (February 2011). "SLAM (CD150) is a multitasking immunoreceptor: from cosignalling to bacterial recognition". Immunology and Cell Biology. 89 (2): 161–3. doi:10.1038/icb.2010.145. PMID 21102539.
- Chatterjee M, Kis-Toth K, Thai TH, Terhorst C, Tsokos GC (May 2011). "SLAMF6-driven co-stimulation of human peripheral T cells is defective in SLE T cells". Autoimmunity. 44 (3): 211–8. doi:10.3109/08916934.2010.530627. PMC 4465387. PMID 21231893.
- Kim JR, Horton NC, Mathew SO, Mathew PA (August 2013). "CS1 (SLAMF7) inhibits production of proinflammatory cytokines by activated monocytes". Inflammation Research. 62 (8): 765–72. doi:10.1007/s00011-013-0632-1. PMID 23695528.
- Wang G, Abadía-Molina AC, Berger SB, Romero X, O'Keeffe MS, Rojas-Barros DI, Aleman M, Liao G, Maganto-García E, Fresno M, Wang N, Detre C, Terhorst C (June 2012). "Cutting edge: Slamf8 is a negative regulator of Nox2 activity in macrophages". Journal of Immunology. 188 (12): 5829–32. doi:10.4049/jimmunol.1102620. PMC 3370125. PMID 22593622.
- Pott S, Kamrani NK, Bourque G, Pettersson S, Liu ET (2012). Jothi R (ed.). "PPARG binding landscapes in macrophages suggest a genome-wide contribution of PU.1 to divergent PPARG binding in human and mouse". PLOS One. 7 (10): e48102. doi:10.1371/journal.pone.0048102. PMC 3485280. PMID 23118933.
- van Driel, Boaz Job; Liao, Gongxian; Engel, Pablo; Terhorst, Cox (2016-01-20). "Responses to Microbial Challenges by SLAMF Receptors". Frontiers in Immunology. 7. doi:10.3389/fimmu.2016.00004. ISSN 1664-3224. PMC 4718992. PMID 26834746.
- Ritchie, David; Colonna, Marco (May 2018). "Mechanisms of Action and Clinical Development of Elotuzumab: Development of Elotuzumab in Myeloma". Clinical and Translational Science. 11 (3): 261–266. doi:10.1111/cts.12532. PMC 5944582. PMID 29272564.
- Sell, Stewart (June 2017). "Cancer immunotherapy: Breakthrough or "deja vu, all over again"?". Tumor Biology. 39 (6): 101042831770776. doi:10.1177/1010428317707764. ISSN 1010-4283. PMID 28639883.