Tetraloop
Tetraloops are a type of four-base hairpin loop motifs in RNA secondary structure that cap many double helices.[2] There are many variants of the tetraloop. The published ones include ANYA,[3][4] CUYG,[5] GNRA,[6] UNAC[7] and UNCG.[8]
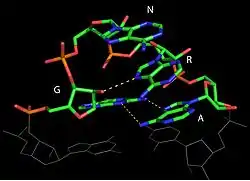
Three types of tetraloops are common in ribosomal RNA: GNRA, UNCG and CUUG, in which the N could be either uracil, adenine, cytosine, or guanine, and the R is either guanine or adenine. These three sequences form stable and conserved tetraloops that play an important role in structural stability and biological function of 16S rRNA.[9]
- GNRA
- The GNRA tetraloop has a guanine-adenine base-pair where the guanine is 5' to the helix and the adenine is 3' to the helix. Tetraloops with the sequence UMAC have essentially the same backbone fold as the GNRA tetraloop,[7] but may be less likely to form tetraloop-receptor interactions. They may therefore be a better choice for closing stems when designing artificial RNAs.
- The presence of the GNRA tetraloop provides an exceptional stability to RNA structure. GNRA occurs 50% more than other tetranucleotides due to their ability to withstand temperatures 4° C higher than other RNA hairpins. This allows them to act as nucleation sites for proper folding of RNA. The rare hydrogen bonds between the first guanine and fourth adenine nucleotide, extensive stacking of nucleotide bases and hydrogen bonds between 2' OH of a ribose sugar and nitrogenous bases makes the tetraloop thermodynamically stable.[10]
- UNCG
- In the UNCG is favorable thermodynamically and structurally due to hydrogen bonding, van der Waals interactions, coulombic interactions and the interactions between the RNA and the solvent. The UNCG tetraloops are more stable than DNA loops with the same sequence. The UUCG tetraloop is the most stable tetraloop.[11] UUCG and GNRA tetraloops make up 70% of all tetraloops in 16S-rRNA .[2]
- CUUG
See also
References
- Cate, J.H., Gooding, A.R., Podell, E., Zhou, K., Golden, B.L., Kundrot, C.E., Cech, T.R., Doudna, J.A. (1996). "Crystal structure of a group I ribozyme domain: principles of RNA packing". Science. 273 (5282): 1676–1685. doi:10.1126/science.273.5282.1678. PMID 8781224.CS1 maint: multiple names: authors list (link)
- Woese, C.R., Winkers, S., Gutell, R.R. (1990). "Architecture of ribosomal RNA: Constraints on the sequence of "tetra-loops"". Proc. Natl. Acad. Sci. USA. 87 (21): 8467–71. doi:10.1073/pnas.87.21.8467. PMC 54977. PMID 2236056.CS1 maint: multiple names: authors list (link)
- Zirbel, CL; Sponer, JE; Sponer, J; Stombaugh, J; Leontis, NB (Aug 2009). "Classification and energetics of the base-phosphate interactions in RNA". Nucleic Acids Research. 37 (15): 4898–918. doi:10.1093/nar/gkp468. PMC 2731888. PMID 19528080.
- Klosterman, PS; Hendrix, DK; Tamura, M; Holbrook, SR; Brenner, SE (2004). "Three-dimensional motifs from the SCOR, structural classification of RNA database: extruded strands, base triples, tetraloops and U-turns". Nucleic Acids Research. 32 (8): 2342–52. doi:10.1093/nar/gkh537. PMC 419439. PMID 15121895.
- Jucker, FM; Pardi, A (Nov 7, 1995). "Solution structure of the CUUG hairpin loop: a novel RNA tetraloop motif". Biochemistry. 34 (44): 14416–27. doi:10.1021/bi00044a019. PMID 7578046.
- Jaeger, L; Michel, F; Westhof, E (Mar 11, 1994). "Involvement of a GNRA tetraloop in long-range RNA tertiary interactions". Journal of Molecular Biology. 236 (5): 1271–6. doi:10.1016/0022-2836(94)90055-8. PMID 7510342.
- Zhao, Q; Huang, HC; Nagaswamy, U; Xia, Y; Gao, X; Fox, GE (Aug 2012). "UNAC tetraloops: to what extent do they mimic GNRA tetraloops?". Biopolymers. 97 (8): 617–28. doi:10.1002/bip.22049. PMID 22605553.
- Molinaro, M; Tinoco I, Jr (Aug 11, 1995). "Use of ultra stable UNCG tetraloop hairpins to fold RNA structures: thermodynamic and spectroscopic applications". Nucleic Acids Research. 23 (15): 3056–63. doi:10.1093/nar/23.15.3056. PMC 307149. PMID 7544890.
- Baumruk, Vladimir; Gouyette, Catherine; Huynh-Dinh, Tam; Sun, Jian-Sheng; Ghomi, Mahmoud (2001-10-01). "Comparison between CUUG and UUCG tetraloops: thermodynamic stability and structural features analyzed by UV absorption and vibrational spectroscopy". Nucleic Acids Research. 29 (19): 4089–4096. doi:10.1093/nar/29.19.4089. ISSN 0305-1048. PMC 60239. PMID 11574692.
- Heus, Hans A.; Pardi, Arthur (1991-01-01). "Structural Features that Give Rise to the Unusual Stability of RNA Hairpins Containing GNRA Loops". Science. 253 (5016): 191–194. doi:10.1126/science.1712983. JSTOR 2878700.
- Antao, V. P.; Lai, S. Y.; Tinoco, I. (1991-11-11). "A thermodynamic study of unusually stable RNA and DNA hairpins". Nucleic Acids Research. 19 (21): 5901–5905. doi:10.1093/nar/19.21.5901. ISSN 0305-1048. PMC 329045. PMID 1719483.
- Hall, Kathleen B. (October 15, 2013). "RNA does the folding dance of twist, turn, stack". Proceedings of the National Academy of Sciences of the United States of America. 110 (42): 16706–7. doi:10.1073/pnas.1316029110. JSTOR 23750643. PMC 3801021. PMID 24072647.
This article is issued from Wikipedia. The text is licensed under Creative Commons - Attribution - Sharealike. Additional terms may apply for the media files.