UBA2
Ubiquitin-like 1-activating enzyme E1B (UBLE1B) also known as SUMO-activating enzyme subunit 2 (SAE2) is an enzyme that in humans is encoded by the UBA2 gene.[5]



Posttranslational modification of proteins by the addition of the small protein SUMO (see SUMO1), or sumoylation, regulates protein structure and intracellular localization. SAE1 and UBA2 form a heterodimer that functions as a SUMO-activating enzyme for the sumoylation of proteins.[5][6]
Structure
DNA
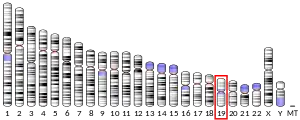
The UBA2 cDNA fragment 2683 bp long and encodes a peptide of 640 amino acids.[6] The predicted protein sequence is more analogous to yeast UBA2 (35% identity) than human UBA3 or E1 (in ubiquitin pathway). The UBA gene is located on chromosome 19.[7]
Protein
Uba2 subunit is 640 aa residues long with a molecular weight of 72 kDa.[8] It consists of three domains: an adenylation domain (containing adenylation active site), a catalytic Cys domain (containing the catalytic Cys173 residue participated in thioester bond formation), and a ubiquitin-like domain. SUMO-1 binds on Uba2 between the catalytic Cys domain and UbL domain.[9]
Mechanism
SUMO activating enzyme (E1, heterodimer of SAE1 and UBA2) catalyzes the reaction of activating SUMO-1 and transferring it to Ubc9 (the only known E2 for SUMOylation). The reaction happens in three steps: adenylation, thioester bond formation, and SUMO transfer to E2. First, the carboxyl group of SUMO C-terminal glycine residue attacks ATP, forming SUMO-AMP and pyrophosphate. Next, the thiol group of a catalytic cysteine in the UBA2 active site attacks SUMO-AMP, forming a high energy thioester bond between UBA2 and the C-terminal glycine of SUMO and releasing AMP. Finally, SUMO is transferred to an E2 cysteine, forming another thioester bond.[9][10][11]
Function
Ubiquitin tag has a well understood role of directing protein towards degradation by proteasome.[12] The role SUMO molecules play are more complicated and much less well understood. SUMOylation consequences include altering substrate affinity for other proteins or with DNA, changing substrate localization, and blocking ubiquitin binding (which prevents substrate degradation). For some proteins, SUMOylation doesn’t seem to have a function.[10][13]
NF-kB
Transcription factor NF-kB in unstimulated cells is inactivated by IkBa inhibitor protein binding. The activation of NF-kB is achieved by ubiquitination and subsequent degradation of IkBa. SUMOylation of IkBa has a strong inhibitory effect on NF-kB-dependent transcription. This may be a mechanism for cell to regulate the number of NF-kB available for transcriptional activation.[14]
p53
Transcription factor p53 is a tumor suppressor acting by activating genes involved in cell cycle regulation and apoptosis. Its level is regulated by mdm2-dependent ubiquitination. SUMOylation of p53 (at a distinct lysine residue from ubiquitin modification sites) prevents proteasome degradation and acts as an additional regulator to p53 response.[15]
Expression and regulation
Studies of yeast budding and fission have revealed that SUMOylation may be important in cell cycle regulation.[16] During a cell cycle, the UBA2 concentration doesn't undergo substantial change while SAE1 level shows dramatic fluctuation, suggesting regulation of SAE1 expression rather than UBA2 might be a way for cell to regulate SUMOylation. However, at time points when SAE1 levels are low, little evidence of UBA2-containing protein complexes are found other than SAE1-UBA2 heterodimer. One possible explanation would be that these complexes exist only for a short period of time, thus not obvious in cell extracts. UBA2 expression is found in most organs including the brain, lung and heart, indicating probable existence of SUMOylation pathway in these organs. An elevated level of UBA2 (as well as all other enzyme components of the pathway) is found in testis, suggesting possible role for UBA2 in meiosis or spermatogenesis. Inside the nucleus, UBA2 is distributed throughout nuclei but not found in nucleoli, suggesting SUMOylation may occur primarily in nuclei. Cytoplasmic existence of SAE 1 and UBA2 is also possible and is responsible for conjugation of cytoplasmic substrates.[17]
Model organisms
Mouse
| Characteristic | Phenotype |
|---|---|
| Homozygote viability | Abnormal |
| Recessive lethal study | Abnormal |
| Body weight | Normal |
| Anxiety | Normal |
| Neurological assessment | Normal |
| Grip strength | Normal |
| Hot plate | Normal |
| Dysmorphology | Normal |
| Indirect calorimetry | Normal |
| Glucose tolerance test | Normal |
| Auditory brainstem response | Normal |
| DEXA | Abnormal[18] |
| Radiography | Abnormal[19] |
| Body temperature | Normal |
| Eye morphology | Normal |
| Clinical chemistry | Normal |
| Plasma immunoglobulins | Normal |
| Haematology | Normal |
| Peripheral blood lymphocytes | Normal |
| Micronucleus test | Normal |
| Heart weight | Normal |
| Skin Histopathology | Normal |
| Brain histopathology | Normal |
| Salmonella infection | Normal[20] |
| Citrobacter infection | Normal[21] |
| All tests and analysis from[22][23] |
Model organisms have been used in the study of UBA2 function. A conditional knockout mouse line, called Uba2tm1a(KOMP)Wtsi[24][25] was generated as part of the International Knockout Mouse Consortium program — a high-throughput mutagenesis project to generate and distribute animal models of disease to interested scientists — at the Wellcome Trust Sanger Institute.[26][27][28]
Male and female animals underwent a standardized phenotypic screen to determine the effects of deletion.[22][29] Twenty five tests were carried out on mutant mice and four significant abnormalities were observed.[22] No homozygous mutant embryos were identified during gestation, and therefore none survived until weaning. The remaining tests were carried out on heterozygous mutant adult mice. Females were found to have a decreased body length by DEXA, while animals of both sex had a decreased number of lumbar and sacral vertebrae in X-rays.[22]
Drosophila
The coding region of drosophila UBA2 homologue dUBA2 gene is 2.3 kb long and contains 2 introns (53 and 52 bp). The predicted protein has 766 amino acid residues and weighs 84 kDa. The protein has an overall identity of 47% to hUBA2 and 31% to yeast UBA2. There are also several regions of complete identity between the three homologous proteins. The C-terminal region of the coding sequence also contains a putative nuclear localization sequence.[30]
Interactions
SAE2 has been shown to interact with
References
- GRCh38: Ensembl release 89: ENSG00000126261 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000052997 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Entrez Gene: ubiquitin-like modifier activating enzyme 2". Retrieved 2011-08-30.
- Okuma T, Honda R, Ichikawa G, Tsumagari N, Yasuda H (January 1999). "In vitro SUMO-1 modification requires two enzymatic steps, E1 and E2". Biochem. Biophys. Res. Commun. 254 (3): 693–8. doi:10.1006/bbrc.1998.9995. PMID 9920803.
- Gong L, Li B, Millas S, Yeh ET (April 1999). "Molecular cloning and characterization of human AOS1 and UBA2, components of the sentrin-activating enzyme complex". FEBS Lett. 448 (1): 185–9. doi:10.1016/S0014-5793(99)00367-1. PMID 10217437. S2CID 7756078.
- Desterro JM, Rodriguez MS, Kemp GD, Hay RT (April 1999). "Identification of the enzyme required for activation of the small ubiquitin-like protein SUMO-1". J. Biol. Chem. 274 (15): 10618–24. doi:10.1074/jbc.274.15.10618. PMID 10187858.
- Lois LM, Lima CD (February 2005). "Structures of the SUMO E1 provide mechanistic insights into SUMO activation and E2 recruitment to E1". EMBO J. 24 (3): 439–51. doi:10.1038/sj.emboj.7600552. PMC 548657. PMID 15660128.
- Johnson ES (2004). "Protein modification by SUMO". Annu. Rev. Biochem. 73: 355–82. doi:10.1146/annurev.biochem.73.011303.074118. PMID 15189146.
- Walden H, Podgorski MS, Schulman BA (March 2003). "Insights into the ubiquitin transfer cascade from the structure of the activating enzyme for NEDD8". Nature. 422 (6929): 330–4. Bibcode:2003Natur.422..330W. doi:10.1038/nature01456. PMID 12646924. S2CID 4370095.
- Müller S, Hoege C, Pyrowolakis G, Jentsch S (March 2001). "SUMO, ubiquitin's mysterious cousin". Nat. Rev. Mol. Cell Biol. 2 (3): 202–10. doi:10.1038/35056591. PMID 11265250. S2CID 5137043.
- Hochstrasser M (October 2001). "SP-RING for SUMO: new functions bloom for a ubiquitin-like protein". Cell. 107 (1): 5–8. doi:10.1016/S0092-8674(01)00519-0. PMID 11595179. S2CID 8021565.
- Desterro JM, Rodriguez MS, Hay RT (August 1998). "SUMO-1 modification of IkappaBalpha inhibits NF-kappaB activation". Mol. Cell. 2 (2): 233–9. doi:10.1016/S1097-2765(00)80133-1. PMID 9734360.
- Rodriguez MS, Desterro JM, Lain S, Midgley CA, Lane DP, Hay RT (November 1999). "SUMO-1 modification activates the transcriptional response of p53". EMBO J. 18 (22): 6455–61. doi:10.1093/emboj/18.22.6455. PMC 1171708. PMID 10562557.
- Saitoh H, Sparrow DB, Shiomi T, Pu RT, Nishimoto T, Mohun TJ, Dasso M (January 1998). "Ubc9p and the conjugation of SUMO-1 to RanGAP1 and RanBP2". Curr. Biol. 8 (2): 121–4. doi:10.1016/S0960-9822(98)70044-2. PMID 9427648.
- Azuma Y, Tan SH, Cavenagh MM, Ainsztein AM, Saitoh H, Dasso M (August 2001). "Expression and regulation of the mammalian SUMO-1 E1 enzyme". FASEB J. 15 (10): 1825–7. doi:10.1096/fj.00-0818fje. PMID 11481243. S2CID 40471133.
- "DEXA data for Uba2". Wellcome Trust Sanger Institute.
- "Radiography data for Uba2". Wellcome Trust Sanger Institute.
- "Salmonella infection data for Uba2". Wellcome Trust Sanger Institute.
- "Citrobacter infection data for Uba2". Wellcome Trust Sanger Institute.
- Gerdin AK (2010). "The Sanger Mouse Genetics Programme: High throughput characterisation of knockout mice". Acta Ophthalmologica. 88: 925–7. doi:10.1111/j.1755-3768.2010.4142.x. S2CID 85911512.
- Mouse Resources Portal, Wellcome Trust Sanger Institute.
- "International Knockout Mouse Consortium".
- "Mouse Genome Informatics".
- Skarnes WC, Rosen B, West AP, Koutsourakis M, Bushell W, Iyer V, Mujica AO, Thomas M, Harrow J, Cox T, Jackson D, Severin J, Biggs P, Fu J, Nefedov M, de Jong PJ, Stewart AF, Bradley A (2011). "A conditional knockout resource for the genome-wide study of mouse gene function". Nature. 474 (7351): 337–42. doi:10.1038/nature10163. PMC 3572410. PMID 21677750.
- Dolgin E (2011). "Mouse library set to be knockout". Nature. 474 (7351): 262–3. doi:10.1038/474262a. PMID 21677718.
- Collins FS, Rossant J, Wurst W (2007). "A mouse for all reasons". Cell. 128 (1): 9–13. doi:10.1016/j.cell.2006.12.018. PMID 17218247. S2CID 18872015.
- van der Weyden L, White JK, Adams DJ, Logan DW (2011). "The mouse genetics toolkit: revealing function and mechanism". Genome Biol. 12 (6): 224. doi:10.1186/gb-2011-12-6-224. PMC 3218837. PMID 21722353.
- Donaghue C, Bates H, Cotterill S (March 2001). "Identification and characterisation of the Drosophila homologue of the yeast Uba2 gene". Biochim. Biophys. Acta. 1518 (1–2): 210–4. doi:10.1016/S0167-4781(01)00185-3. PMID 11267682.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (October 2005). "Towards a proteome-scale map of the human protein-protein interaction network". Nature. 437 (7062): 1173–8. Bibcode:2005Natur.437.1173R. doi:10.1038/nature04209. PMID 16189514. S2CID 4427026.
- Ewing RM, Chu P, Elisma F, Li H, Taylor P, Climie S, McBroom-Cerajewski L, Robinson MD, O'Connor L, Li M, Taylor R, Dharsee M, Ho Y, Heilbut A, Moore L, Zhang S, Ornatsky O, Bukhman YV, Ethier M, Sheng Y, Vasilescu J, Abu-Farha M, Lambert JP, Duewel HS, Stewart II, Kuehl B, Hogue K, Colwill K, Gladwish K, Muskat B, Kinach R, Adams SL, Moran MF, Morin GB, Topaloglou T, Figeys D (2007). "Large-scale mapping of human protein-protein interactions by mass spectrometry". Mol. Syst. Biol. 3 (1): 89. doi:10.1038/msb4100134. PMC 1847948. PMID 17353931.
- Tatham MH, Kim S, Yu B, Jaffray E, Song J, Zheng J, Rodriguez MS, Hay RT, Chen Y (August 2003). "Role of an N-terminal site of Ubc9 in SUMO-1, -2, and -3 binding and conjugation". Biochemistry. 42 (33): 9959–69. doi:10.1021/bi0345283. PMID 12924945.
- Knipscheer P, Flotho A, Klug H, Olsen JV, van Dijk WJ, Fish A, Johnson ES, Mann M, Sixma TK, Pichler A (August 2008). "Ubc9 sumoylation regulates SUMO target discrimination". Mol. Cell. 31 (3): 371–82. doi:10.1016/j.molcel.2008.05.022. PMID 18691969.
Further reading
- Pichler A, Gast A, Seeler JS, Dejean A, Melchior F (2002). "The nucleoporin RanBP2 has SUMO1 E3 ligase activity". Cell. 108 (1): 109–20. doi:10.1016/S0092-8674(01)00633-X. PMID 11792325. S2CID 14656901.
- Boggio R, Colombo R, Hay RT, Draetta GF, Chiocca S (2004). "A mechanism for inhibiting the SUMO pathway". Mol. Cell. 16 (4): 549–61. doi:10.1016/j.molcel.2004.11.007. PMID 15546615.
- Stelzl U, Worm U, Lalowski M, Haenig C, Brembeck FH, Goehler H, Stroedicke M, Zenkner M, Schoenherr A, Koeppen S, Timm J, Mintzlaff S, Abraham C, Bock N, Kietzmann S, Goedde A, Toksöz E, Droege A, Krobitsch S, Korn B, Birchmeier W, Lehrach H, Wanker EE (2005). "A human protein-protein interaction network: a resource for annotating the proteome". Cell. 122 (6): 957–68. doi:10.1016/j.cell.2005.08.029. hdl:11858/00-001M-0000-0010-8592-0. PMID 16169070. S2CID 8235923.
- Li T, Santockyte R, Shen RF, Tekle E, Wang G, Yang DC, Chock PB (2006). "A general approach for investigating enzymatic pathways and substrates for ubiquitin-like modifiers". Arch. Biochem. Biophys. 453 (1): 70–4. doi:10.1016/j.abb.2006.03.002. PMID 16620772.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (2005). "Towards a proteome-scale map of the human protein-protein interaction network". Nature. 437 (7062): 1173–8. Bibcode:2005Natur.437.1173R. doi:10.1038/nature04209. PMID 16189514. S2CID 4427026.
- Knipscheer P, Flotho A, Klug H, Olsen JV, van Dijk WJ, Fish A, Johnson ES, Mann M, Sixma TK, Pichler A (2008). "Ubc9 sumoylation regulates SUMO target discrimination". Mol. Cell. 31 (3): 371–82. doi:10.1016/j.molcel.2008.05.022. PMID 18691969.
- Gilbreth RN, Truong K, Madu I, Koide A, Wojcik JB, Li NS, Piccirilli JA, Chen Y, Koide S (2011). "Isoform-specific monobody inhibitors of small ubiquitin-related modifiers engineered using structure-guided library design". Proc. Natl. Acad. Sci. U.S.A. 108 (19): 7751–6. Bibcode:2011PNAS..108.7751G. doi:10.1073/pnas.1102294108. PMC 3093456. PMID 21518904.
- Moutty MC, Sakin V, Melchior F (2011). "Importin α/β mediates nuclear import of individual SUMO E1 subunits and of the holo-enzyme". Mol. Biol. Cell. 22 (5): 652–60. doi:10.1091/mbc.E10-05-0461. PMC 3046061. PMID 21209321.
- Bossis G, Melchior F (2006). "Regulation of SUMOylation by reversible oxidation of SUMO conjugating enzymes". Mol. Cell. 21 (3): 349–57. doi:10.1016/j.molcel.2005.12.019. PMID 16455490.
- Lois LM, Lima CD (2005). "Structures of the SUMO E1 provide mechanistic insights into SUMO activation and E2 recruitment to E1". EMBO J. 24 (3): 439–51. doi:10.1038/sj.emboj.7600552. PMC 548657. PMID 15660128.
- Azuma Y, Tan SH, Cavenagh MM, Ainsztein AM, Saitoh H, Dasso M (2001). "Expression and regulation of the mammalian SUMO-1 E1 enzyme". FASEB J. 15 (10): 1825–7. doi:10.1096/fj.00-0818fje. PMID 11481243. S2CID 40471133.
- Lemos TA, Kobarg J (2006). "CGI-55 interacts with nuclear proteins and co-localizes to p80-coilin positive-coiled bodies in the nucleus". Cell Biochem. Biophys. 44 (3): 463–74. doi:10.1385/CBB:44:3:463. PMID 16679534. S2CID 24544907.
- Li L, Liang D, Li JY, Zhao RY (2008). "APOBEC3G-UBA2 fusion as a potential strategy for stable expression of APOBEC3G and inhibition of HIV-1 replication". Retrovirology. 5: 72. doi:10.1186/1742-4690-5-72. PMC 2535603. PMID 18680593.
- Tatham MH, Kim S, Yu B, Jaffray E, Song J, Zheng J, Rodriguez MS, Hay RT, Chen Y (2003). "Role of an N-terminal site of Ubc9 in SUMO-1, -2, and -3 binding and conjugation". Biochemistry. 42 (33): 9959–69. doi:10.1021/bi0345283. PMID 12924945.
- Beausoleil SA, Jedrychowski M, Schwartz D, Elias JE, Villén J, Li J, Cohn MA, Cantley LC, Gygi SP (2004). "Large-scale characterization of HeLa cell nuclear phosphoproteins". Proc. Natl. Acad. Sci. U.S.A. 101 (33): 12130–5. Bibcode:2004PNAS..10112130B. doi:10.1073/pnas.0404720101. PMC 514446. PMID 15302935.
External links
- UBA2 human gene location in the UCSC Genome Browser.
- UBA2 human gene details in the UCSC Genome Browser.
PDB gallery | |
|---|---|
|