Ulota
Ulota is a genus of mosses comprising 69 species with a worldwide distribution, though most species are found in the southern hemisphere.
| Ulota | |
|---|---|
_7767.JPG.webp) | |
| Ulota coarctata | |
| Scientific classification | |
| Kingdom: | Plantae |
| Division: | Bryophyta |
| Class: | Bryopsida |
| Subclass: | Bryidae |
| Order: | Orthotrichales |
| Family: | Orthotrichaceae |
| Genus: | Ulota Mohr |
.jpg.webp)
They have commonly been grouped within the genus Orthotrichum, though recently they have been placed in their own genus. Their phylogeny and taxonomy have been an area of debate ever since they were placed in their own genus, though it has been largely accepted to place Ulota as a genus of their own. They can be found growing on trees close to the coast, though some species are also found on rocks. Their shoots form small tufts to large mats across habitats, often intertwined with other mosses, including Orthotrichum species.
Ulota reproduce using sexual structures, sporangium (plural: sporangia), that are terminal on the shoot. The calyptra covering the developing sporangium can be hairy or not hairy depending on the species, but the hairs extend from the sporangium base to the apex as opposed to the calyptra hairs of Polytrichum mosses which extend from the apex to the base.
Taxonomy
Due to morphological similarities, this genus has often been taxonomically clustered with the genus Orthotrichum, and has also been considered as a subgenus of Orthotrichum.[1][2] Often, these two genus are found growing together. Mohr first segregated Ulota from Orthotrichum in 1806, though the segregation has been debated. The two distinguishing features were the splitting of the calyptra at the base and the curled edges of the leaves. However, some Orthotrichum species also have calyptrae which split at the base.[3] Molecular data has made some progress as to differentiating the taxa, though certain morphological features can be distinguishing.[4] Ulota have superficial stomata, whereas Orthotrichum have cryptoporous stomata.[5] Other distinguishing features of Ulota include being monoicous, and the absence of asexual reproductive structures.[5] There is one dioicous species U. phyllantha, and according to molecular studies it is closer related to other Ulota species than Orthotrichum species which can be dioicous.[6] The hyaline cells at the base of the leaf margin also helps to distinguish between Orthotrichum and most Ulota species.[3]
Recent genetic analysis of the mitochondrial genomes of species within Orthotrichum suggests the presence of seven genera within the taxon.[7] The seven genera include: the genus Orthotrichum, which groups together the three previous subgenera Callistoma, Pulchella, and Orthotrichum;[7] the genus Leratia, comprising the previous subgenus Exiguifolium;[8] the genus Lewinskya, which includes the two previous subgenera Phaneroporum and Gymnoporus;[9] the genus Pulvigera, comprising only one species, Orthotrichum lyelli;[5] the genus Nyholmiella, composed of the previous subgenus Orthophyllum;[6] the genus Plenogemma, composed of the single species U. phyllantha;[5] and the genus Ulota, comprising all previous species classified as Ulota.[7]
Species list
There are currently 69 recognized species of Ulota, over half of which occur only in the southern hemisphere. This number also includes U. phyllantha which has been proposed to reside within its own genus.[2][5]
- U. angusti-limbata E.B. Bartram
- U. angustissima Müller Hal.
- U. aurantiaca Dusen ex Malta
- U. barclayi Mitt.
- U. bellii Malta
- U. billbuckii Garilleti, Mazimpaka & F. Lara
- U. breviseta Malta
- U. bruchii Hornsch.
- U. calvescens Wilson
- U. carinata Mitt.
- U. cervina Hoe & H.A. Crum
- U. coarctata (P. Beauv.) Hammar
- U. cochleata Venturi
- U. crispa (Hedw.) Bridel
- U. crispula Bruch
- U. curvifolia (Wahlenb.) Lilj.
- U. delicata Q.-H. Wang & Y. Jia
- U. dixonii Malta
- U. drummondii (Hook. & Grev.) Brid.
- U. ecklonii Hornsch. A. Jaeger
- U. eurystoma Nog.
- U. fernandeziana Malta
- U. fuegiana Mitt.
- U. fulva Brid.
- U. germana (Mont.) Mitt.
- U. gigantospora F. Lara, Caparrós & Garilleti
- U. glabella Mitt.
- U. gymnostoma Guo, Enroth & Virtanen
- U. hutchinsiae var. hutchinsiae]] (Sm.) Hammar
- U. hutchinsiae var. rufescens]] (E. Britton) Dixon
- U. japonica (Sull. & Lesq.) Mitt.
- U. larrainii Garilleti, Mazimpaka & F. Lara
- U. laticiliata Malta
- U. latisegmenta Q.-H. Wang & Y. Jia
- U. lativentrosa Müll. Hal. ex Malta
- U. longifolia Dixon & Sakurai
- U. lutea var. lutea (Hook. f. & Wilson) Mitt.
- U. lutea var. robusta Dixon ex Malta
- U. luteola (Hook. f. & Wilson) Mitt.
- U. macrocalycina Mitt.
- U. macrodontia Dusén ex Malta
- U. macrospora Baur & Warnst.
- U. magellanica (Mont.) A. Jaeger
- U. megalospora Venturi
- U. membranata Malta
- U. morrisonensis Horik.
- U. novae-seelandiaei Sainsbury
- U. novo-guinensis Bartram
- U. obtusiuscula Müll. Hal. & Kindb.
- U. perbreviseta Dixon & Sakurai
- U. perichaetialis (Sainsbury) Goffinet
- U. phyllantha Brid.
- U. phyllanthoides (Müll. Hal.) Kindb.
- U. pusilla Malta
- U. pycnophylla Dusén ex Malta
- U. pygmaeothecia (Müll. Hal.) Kindb.
- U. rehmannii Jur.
- U. reptans Mitt.
- U. rhytiore (B.H. Allen) F. Lara, Garilleti, Albertos & Mazimpaka
- U. robusta Mitt.
- U. rubella E.B. Bartram
- U. rufula (Mitt.) A. Jaeger
- U. schmidii (Müll. Hal.) Mitt.
- U. splendida E.B. Bartram
- U. streptodon Garilleti, Mazimpaka & F. Lara
- U. ventricosa (Müll. Hal.) Malta
- U. viridis Venturi
- U. yakushimensis Z. Iwatsuki
- U. yunnanensis F. Lara, Caparrós & Garilleti
Description
_8609.JPG.webp)
_0442.JPG.webp)
Ulota mosses tend to grow in a tuft on the substrate, with the shoots growing in an acrocarpous fashion with the shoots standing up.[10] The tufts can vary in colour, ranging from a deep green at the top of the tuft to a darker brown/red at the base of the cushion.[4] The oblong to linear-lanceolate leaves are attached to an erect stem, and can be slightly flexed to fully curled when dry.[11][5] The genus is characterized by the slightly to fully curled leaves when dry. The leaves have a costa, or midrib, which runs the entire length of the leaf.[5] The leaves have hyaline cells at the basal margins of the leaves, which are typically square or rectangular shaped.[11] The leaf base can be expanded or gradually tapered as it connects to the stem.[12] At the tip of the leaves, there are cells with very thick cell walls.[13] At maturity, the sporangia can be ribbed or unribbed.[4] The sporangia sit on top of a seta, which holds it above the gametophyte. Between species of Ulota, there is variation in the length of the seta which can be a useful trait in classifying species.[12] The sporangia have peristome teeth are diplolepidous, with 8 to 16 exostome teeth and 8 endostome teeth.[11][5] The sporangia sit on top of a seta which attaches to the gametophyte at the apex of the shoot.[3] The calyptra covers the developing sporangium and is typically conic, split at the base, and can be naked or hairy.[5] The rhizoids are placed at the base of them stem, occasionally appearing along the stem, and are smooth.[4]
Habitat and distribution
Ulota species occur typically in temperate climates, growing as an epiphyte on tree trunks. Some species have also been known to extend into the tropics and subarctic environments.[11] Multiple species can grow together in a habitat, often intertwined with each other.[14] The genus is typically found in temperate, humid, oceanic coasts, though some species are also found in montane environments in the tropics and subtropics.[4] Their habitat is found to be heavily determined by the available substrate, but also on the dispersal ability of the species and the dispersal obstacles that are present.[15] The species which tend to be correlated with the presence of Ulota species include: Salix caprea, Populus tremula, Fraxinus excelsior, Sorbus aucuparia, and Acer pseudoplatanus.[15] They can also be found growing on rocks, though this is reportedly less common.[4] Two species, U. crispa and U. bruchii, tend to be found together, though U. crispa tends to be more common along the coast.[16]
Within their distribution, four areas have the highest rates of endemism for Ulota species: Papua New Guinea (75%), Subtropical China (77.8%), Southern Australasia (78.6%), and South America (81.8%). South America also has the highest richness of Ulota species with 22 recorded species. The next richest area is Australasia with 14 recognized species.[2]
Lifecycle
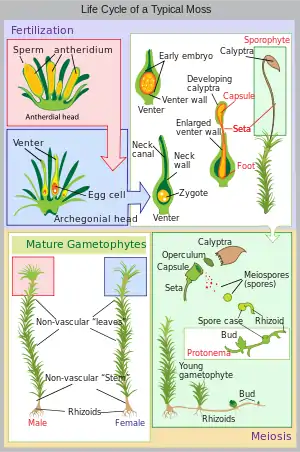
Like other bryophytes, show an alternation of generations.[17] The vegetative gametophyte generation is haploid, and is dominant for most of the life history.[18] Most plants are dioicous except for U. phyllantha, which is monoicous.[6] Asexual reproduction is uncommon, except in U. phyllantha, in which asexual propagules can commonly be found at the leaf apex.[19]
Sexual reproduction involves the transfer of sperm from the antheridium (plural: antheridia) via water. Drops of water carrying sperm allow for the fertilization of the egg housed within the archegonium (plural: archegonia). The heads of the archegonia have a mucilage which attracts the sperm, and the degeneration of the neck canal allows for the passage of sperm to the egg, resulting in fertilization. The fertilized egg develops into a sporophyte, which is diploid, and protected by a remnant of the archegonium now known as the calyptra. Inside the developing sporangium, haploid spores are generated via meiosis. The seta raises the sporangium into the air, and once the sporangium is mature, the operculum falls off and the spores are released into the wind. The peristome teeth are hygroscopic and move in relation to the moisture content of the air, which allows for spores to be released over a longer period of time.[17] Once mature, the sporangia are typically 8-ribbed, though in some species the ribs are only clearly visible right at the base of the peristome mouth.[4]
Spores can be unicellular, although there five known species in which multicellular spores have been identified. Endosporic germination is the process in which mitosis occurs within the sporeling before the spore is released, and once released, it is considered a multicellular spore.[14] This kind of germination is known in all three lineages of bryophytes: liverworts, hornworts, and mosses.[20] Once the spores have landed on suitable substrata, protonematal threads will begin to grow, followed by rhizoids which will help attach the developing plant to the substrate.[14] The leafy shoot develops soon after, reaching upwards towards light.[14]
Conservation
_5284.jpg.webp)
Of the 69 recognized species, three of the species have been assessed by the IUCN.[21] Native to northwestern Europe and Serbia, U. calvescens shows an increasing population trend and is listed "least concerned". There is one protected area within its range, and it is listed as vulnerable in Portugal, Spain, and Serbia, and data deficient in Norway.[22] U. bruchii is found throughout northern and western Europe, with a few pockets in eastern Europe and is listed as "least concerned".[23] U. macrospora exists in a small area within central Europe, and is listed as "endangered".[24] It has a small population size and the current threats it faces includes a decline in habitat area and lowering of habitat quality.[24] Declines in air quality have also been correlated with declines in the size of the tufts of this genus found on trees in urban areas.[25]
Uses
Ulota crispa and U. megalospora have both been found to be intolerant of high levels of sulfur dioxide, which may indicate both species' use as an air quality indicator.[26][25]
References
- Mitten, William (1869). Musci Austro-Americani: Sive Enumeratio Muscorum Omnium Austro-Americanorum Mihi Hucusque Cognitorum, Eorum Praecipue in Terris Amazonicis Andinisque Ricardo Spruceo Lectorum. Longmans, Green, Reader, & Dyer.
- Garilleti, Ricardo; Mazimpaka, Vicente; Lara, Francisco (2015). "Ulota larrainii (Orthotrichoideae, Orthotrichaceae, Bryophyta) a new species from Chile, with comments on the worldwide diversification of the genus". Phytotaxa. 217 (2): 133–144. doi:10.11646/phytotaxa.217.2.3 – via Biotaxa.
- Wang, Qing-Hua; Jia, Yu (November 2012). "A taxonomic revision of the Asian species of Ulota Mohr (Orthotrichaceae)". The Bryologist. 115 (3): 412–443. doi:10.1639/0007-2745-115.3.412. ISSN 0007-2745. S2CID 86387122.
- Caparrós, Rut; Lara, Francisco; Draper, Isabel; Mazimpaka, Vicente; Garilleti, Ricardo (2016-04-01). "Integrative taxonomy sheds light on an old problem: the Ulota crispa complex (Orthotrichaceae, Musci)". Botanical Journal of the Linnean Society. 180 (4): 427–451. doi:10.1111/boj.12397. ISSN 0024-4074.
- Plášek, Vítězslav; Sawicki, Jakub; Ochyra, Ryszard; Szczecińska, Monika; Kulik, Tomasz (2015). "New taxonomical arrangement of the traditionally conceived genera Orthotrichum and Ulota (Orthotrichaceae, Bryophyta)". Acta Musei Silesiae, Scientiae Naturales. 64 (2): 169–174. doi:10.1515/cszma-2015-0024. ISSN 2336-3207.
- Sawicki, Jakub; Plášek, Vítězslav; Szczecińska, Monika (2010). "Molecular studies resolve Nyholmiella (Orthotrichaceae) as a separate genus". Journal of Systematics and Evolution. 48 (3): 183–194. doi:10.1111/j.1759-6831.2010.00076.x. ISSN 1759-6831. S2CID 84318916.
- Sawicki, Jakub; Plášek, Vítězslav; Ochyra, Ryszard; Szczecińska, Monika; Ślipiko, Monika; Myszczyński, Kamil; Kulik, Tomasz (2017-06-30). "Mitogenomic analyses support the recent division of the genus Orthotrichum (Orthotrichaceae, Bryophyta)". Scientific Reports. 7 (1): 4408. Bibcode:2017NatSR...7.4408S. doi:10.1038/s41598-017-04833-z. ISSN 2045-2322. PMC 5493672. PMID 28667304.
- Goffinet, B.; Shaw, A. J.; Cox, C. J.; Wickett, N. J.; Boles, S. (2004). "Phylogenetic inferences in the Orthotrichoideae (Orthotrichaceae: Bryophyta) based on variation in four loci from all genomes". Monographs in Systematic Botany from the Missouri Botanical Garden. 98: 270–289.
- Lara, Francisco; Garilleti, Ricardo; Goffinet, Bernard; Draper, Isabel; Medina, Rafael; Vigalondo, Beatriz; Mazimpaka, Vicente (2016). "Lewinskya, a New Genus to Accommodate the Phaneroporous and Monoicous Taxa of Orthotrichum (Bryophyta, Orthotrichaceae)". Cryptogamie, Bryologie. 37 (4): 361–382. doi:10.7872/cryb/v37.iss4.2016.361. ISSN 1290-0796. S2CID 88561982.
- Caparrós, Rut; Lara, Francisco; Long, David G.; Mazimpaka, Vicente; Garilleti, Ricardo (2011-09-01). "Two new species of Ulota (Orthotrichaceae, Bryopsida) with multicellular spores, from the Hengduan Mountains, Southwestern China". Journal of Bryology. 33 (3): 210–220. doi:10.1179/1743282011Y.0000000008. ISSN 0373-6687. S2CID 84279281.
- Wang, Qinghua; Jia, Yu (2016-04-01). "A taxonomic revision of Ulota Mohr (Orthotrichaceae) in South and Central America". Plant Diversity. 38 (2): 65–88. doi:10.1016/j.pld.2016.03.003. ISSN 2468-2659. PMC 6112220. PMID 30159451.
- Blockeel, Tom; Bosanquet, Sam (May 2017). "The Ulota crispa group in Britain and Ireland, with notes on other species of the genus" (PDF). Field Bryology. 117: 8–19 – via British Bryological Society.
- Vitt, Dale; Koponen, Timo; Norris, Daniel (1993). "Bryophyte flora of the Huon Peninsula, Papua New Guinea. LIII". Acta Botanica Fennica. 148: 5–25 – via ResearchGate.
- Garilleti, Ricardo; Mazimpaka, Vicente; Lara, Francisco (December 2012). "New Ulota species with multicellular spores from southern South America". The Bryologist. 115 (4): 585–600. doi:10.1639/0007-2745-115.4.585. ISSN 0007-2745. S2CID 86367474.
- Heegaard, Einar; Hangelbroek, Helen H. (1999). "The Distribution of Ulota crispa at a Local Scale in Relation to Both Dispersal- and Habitat-Related Factors". Lindbergia. 24 (2): 65–74. ISSN 0105-0761. JSTOR 20150006.
- Albertos, B.; Lara, F.; Garilleti, R.; Mazimpaka, V. (2000). "Distribution and abundance of corticolous species of the genus Ulota (Orthotrichaceae, Musci) in the NW Iberian Peninsula". Nova Hedwigia. 70: 461–470. hdl:11268/5759.
- McMillin, John (1966). "A Laboratory Investigation of the Mosses". The American Biology Teacher. 28 (10): 814–817. doi:10.2307/4441553. ISSN 0002-7685. JSTOR 4441553.
- During, Heinjo J. (1979). "Life Strategies of Bryophytes: A Preliminary Review". Lindbergia. 5 (1): 2–18. ISSN 0105-0761. JSTOR 20149317.
- Seppelt, R. D. (1978-03-01). "Studies on the bryoflora of Macquarie Island". New Zealand Journal of Botany. 16 (1): 21–23. doi:10.1080/0028825X.1978.10429654. ISSN 0028-825X.
- Schuette, Scott; Renzaglia, Karen S. (2010). "Development of multicellular spores in the hornwort genus Dendroceros (Dendrocerotaceae, Anthocerotophyta) and the occurrence of endospory in Bryophytes". Nova Hedwigia. 91 (3): 301–316. doi:10.1127/0029-5035/2010/0091-0301.
- "The IUCN Red List of Threatened Species". IUCN Red List of Threatened Species. Retrieved 2020-04-05.
- Consultant), Nick Hodgetts (Europe LIFE project-Freelance (2017-05-20). "IUCN Red List of Threatened Species: Balding Pincushion". IUCN Red List of Threatened Species. Retrieved 2020-04-05.
- Consultant), Nick Hodgetts (Europe LIFE project-Freelance (2017-05-20). "IUCN Red List of Threatened Species: Bruch's pincushion". IUCN Red List of Threatened Species. Retrieved 2020-04-05.
- Vana, Jiri; Norbert Schnyder (University of Zürich, Switzerland); Consultant), Nick Hodgetts (Europe LIFE project-Freelance; History), Irene Bisang (Swedish Museum of Natural; Baudraz, Maude; Kučera, Jan; Ştefănuţ, Sorin; Lars Hedenäs (Natural History Museum, Stockholm); Steffen Caspari (Zentrum für Biodokumentation, Schiffweiler (2017-01-04). "IUCN Red List of Threatened Species: Ulota macrospora". IUCN Red List of Threatened Species. Retrieved 2020-04-05.
- Hallingbäck, Tomas (1992-01-01). "The effect of air pollution on mosses in southern Sweden". Biological Conservation. Endangered Bryophytes in Europe-Causes and Conservation. 59 (2): 163–170. doi:10.1016/0006-3207(92)90577-A. ISSN 0006-3207.
- Ireland, Robert R. (2004). "Review of Grimmias of the World". The Bryologist. 107 (2): 274–276. doi:10.1639/0007-2745(2004)107[0275:R]2.0.CO;2. ISSN 0007-2745. JSTOR 3244857.