XB130
XB130 (also known as AFAP1L2) is a cytosolic adaptor protein and signal transduction mediator. XB130 regulates cell proliferation, cell survival, cell motility and gene expression. XB130 is highly similar to AFAP and is thus known as actin filament associated protein 1-like 2 (AFAP1L2). XB130 is a substrate and regulator of multiple tyrosine kinase-mediated signaling. XB130 is highly expressed in the thyroid and spleen.


Molecular structure
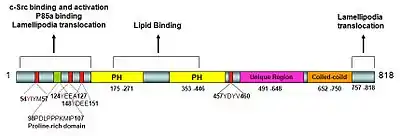
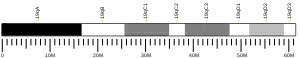
The XB130 gene is located on human chromosome 10q25.3 and encodes an 818 amino acid protein. It has a molecular weight of approximately 130 kDa and is structurally similar to actin-filament-associated protein (AFAP) and is thus known as AFAP1L2.[5] Several tyrosine phosphorylation sites and a proline rich sequence are included in the N-terminal region of XB130, which allows it to interact and activate c-Src-containing proteins, as well as bind to p85α of PI3K. Two pleckstrin-homology domains are located in the middle portion, giving XB130 its lipid-binding ability. The C-terminal region contains a coiled-coil domain, which shares partial similarity with AFAP's leucine zipper domain.[6] Both the C-terminal and N-terminal regions of XB130 are required for XB130's role in its translocation to the lamellipodia.[7] Despite XB130's structural similarity to AFAP, XB130 does not behave like an actin filament-associated protein. The actin-binding site present in AFAP is only partially present in XB130.
Function
Role in cell cycle and survival
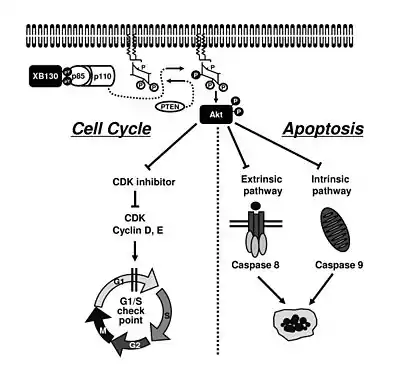
XB130 has been demonstrated to play a role in cell proliferation and survival through the regulation of the PI3K/Akt signaling pathway. When tyrosine phosphorylated, XB130 has the ability to interact with the p85ɑ subunit of PI3K through its SH2 domains.[7] This interaction leads to the subsequent activation of Akt, cell proliferation, and cell survival. Activated Akt promotes cell survival and cell cycle progression by phosphorylating and inactivating p21Cip1/WAF1, p27Kip1, and GSK3β, as well as inhibits apoptosis by preventing the cleavage of caspase-8 and caspase-9, which are involved in the extrinsic and intrinsic pathways of cell death, respectively.[8]> Alternatively, when the expression of XB130 is suppressed in vitro, Akt phosphorylation and therefore activation becomes significantly reduced. This, in turn, leads to cell cycle arrest at G1/S phase and accelerated apoptosis.[9]
Role in cell motility and invasion
During cytoskeletal rearrangement, a process required for cell motility, XB130 translocates to the cell periphery. XB130 exhibits a high affinity for peripheral F-actin structures, such as the lamellipodium. The translocation of XB130 to the cell periphery is particularly important in its potential to influence cell migration and metastasis.[7]
Role in gene expression
The level of XB130 expression influences the expression of multiple genes related to cell proliferation and survival,[10] and microRNAs miR-33a, 149a, and 193a-3p, all of which exhibit tumor suppressive function in thyroid cancer cells.[11]
Role in inflammation
XB130 mediated c-Src binding and activation increases Interleukin-8 (IL-8), a chemokine produce by lung epithelial cells, which contains AP-1 and SRE transcription factor binding sites. These binding sites can be activated by the downregulation of XB130 expression and lead to a decrease in IL-8 production in lung cells.[5]
Interactions
XB130 (gene) has been known to interact with
- SH2 domain of Src
- SH3 domain of Src
- c-Src
- p85a subunit of PI3K
- RET/PTC
- GTPase-activating proteins (GAP)
- Phospholipase C-gamma (PLC-γ)
Clinical significance
Adaptor proteins play an important role as molecular scaffolds to mediate the transport and interaction of various proteins and is therefore highly involved in signal transduction.[5] Deregulation of adaptor proteins is highly related to the abnormality of cellular functions and many adaptor proteins are frequently overexpressed in cancers. Clinical studies on the expression level and pattern of XB130 in various human tumors demonstrate that XB130 expression is regulated in thyroid [10] and gastrointestinal cancer[12][13][14] and soft tissue tumors.[15] Expression level of XB130 was significantly higher in normal and benign lesion than that of papillary and anaplastic/insular carcinoma.[10] Through the studies on many gastrointestinal cancers, the oncogenic roles of XB130 was shown. XB130 expression is significantly correlated with the survival time and disease-free period in gastric cancer patients.[12] XB130 was identified as a potential colorectal cancer markers.[13] XB130 protein level was elevated in human esophageal squamous carcinoma.[14] In addition, XB130 was selected as one of six highly expressed genes related to local aggressiveness of soft tissue tumors in a set of 102 representative tumor samples.[15] These findings suggest that XB130 may be involved in tumorigenesis and that XB130 is a potential diagnostic biomarker and therapeutic target for cancer.
Discovery
This adaptor protein was discovered during molecular cloning of human actin filament associated protein (AFAP1) in the Latner Thoracic Surgery Research Laboratories Toronto, Ontario, Canada. The molecule is named XB130 after lead technician Xiaohui Bai and the molecular mass of the protein. This protein was found to have a high sequence identity to AFAP1, thus its name AFAP1L2.
References
- GRCh38: Ensembl release 89: ENSG00000169129 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000025083 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Zhang R, Zhang J, Wu Q, Meng F, Liu C (2016). "XB130: A novel adaptor protein in cancer signal transduction". Biomedical Reports. 4 (3): 300–306. doi:10.3892/br.2016.588. PMC 4774376. PMID 26998266.
- Snyder BN, Cho Y, Qian Y, Coad JE, Flynn DC, Cunnick JM (2011). "AFAP1L1 is a novel adaptor protein of the AFAP family that interacts with cortactin and localizes to invadosomes". European Journal of Cell Biology. 90 (5): 376–89. doi:10.1016/j.ejcb.2010.11.016. PMC 3085893. PMID 21333378.
- Lodyga M, Bai XH, Kapus A, Liu M (2010). "Adaptor protein XB130 is a Rac-controlled component of lamellipodia that regulates cell motility and invasion". Journal of Cell Science. 123 (Pt 23): 4156–69. doi:10.1242/jcs.071050. PMID 21084565.
- Shiozaki A, Shen-Tu G, Bai X, Iitaka D, De Falco V, Santoro M, Keshavjee S, Liu M (2012). "XB130 mediates cancer cell proliferation and survival through multiple signaling events downstream of Akt". PLOS ONE. 7 (8): e43646. doi:10.1371/journal.pone.0043646. PMC 3426539. PMID 22928011.
- Saini KS, Loi S, de Azambuja E, Metzger-Filho O, Saini ML, Ignatiadis M, Dancey JE, Piccart-Gebhart MJ (2013). "Targeting the PI3K/AKT/mTOR and Raf/MEK/ERK pathways in the treatment of breast cancer". Cancer Treatment Reviews. 39 (8): 935–46. doi:10.1016/j.ctrv.2013.03.009. PMID 23643661.
- Shiozaki A, Lodyga M, Bai XH, Nadesalingam J, Oyaizu T, Winer D, Asa SL, Keshavjee S, Liu M (2011). "XB130, a novel adaptor protein, promotes thyroid tumor growth". The American Journal of Pathology. 178 (1): 391–401. doi:10.1016/j.ajpath.2010.11.024. PMC 3070596. PMID 21224076.
- Takeshita H, Shiozaki A, Bai XH, Iitaka D, Kim H, Yang BB, Keshavjee S, Liu M (2013). "XB130, a new adaptor protein, regulates expression of tumor suppressive microRNAs in cancer cells". PLOS ONE. 8 (3): e59057. doi:10.1371/journal.pone.0059057. PMC 3602428. PMID 23527086.
- Shi M, Huang W, Lin L, Zheng D, Zuo Q, Wang L, Wang N, Wu Y, Liao Y, Liao W (2012). "Silencing of XB130 is associated with both the prognosis and chemosensitivity of gastric cancer". PLOS ONE. 7 (8): e41660. doi:10.1371/journal.pone.0041660. PMC 3426513. PMID 22927913.
- Emaduddin M, Edelmann MJ, Kessler BM, Feller SM (2008). "Odin (ANKS1A) is a Src family kinase target in colorectal cancer cells". Cell Communication and Signaling. 6: 7. doi:10.1186/1478-811X-6-7. PMC 2584000. PMID 18844995.
- Shiozaki A, Kosuga T, Ichikawa D, Komatsu S, Fujiwara H, Okamoto K, Iitaka D, Nakashima S, Shimizu H, Ishimoto T, Kitagawa M, Nakou Y, Kishimoto M, Liu M, Otsuji E (2013). "XB130 as an independent prognostic factor in human esophageal squamous cell carcinoma". Annals of Surgical Oncology. 20 (9): 3140–50. doi:10.1245/s10434-012-2474-4. PMID 22805860. S2CID 10246459.
- Cunha IW, Carvalho KC, Martins WK, Marques SM, Muto NH, Falzoni R, Rocha RM, Aguiar S, Simoes AC, Fahham L, Neves EJ, Soares FA, Reis LF (2010). "Identification of genes associated with local aggressiveness and metastatic behavior in soft tissue tumors". Translational Oncology. 3 (1): 23–32. doi:10.1593/tlo.09166. PMC 2822450. PMID 20165692.