Chlororespiration
Chlororespiration is a respiratory process that takes place within plants. Inside plant cells there is an organelle called the chloroplast which is surrounded by the thylakoid membrane. This membrane contains an enzyme called NAD(P)H dehydrogenase which transfers electrons in a linear chain to oxygen molecules.[1] This electron transport chain (ETC) within the chloroplast also interacts with those in the mitochondria where respiration takes place.[2] Photosynthesis is also a process that Chlororespiration interacts with.[2] If photosynthesis is inhibited by environmental stressors like water deficit, increased heat, and/or increased/decreased light exposure, or even chilling stress then chlororespiration is one of the crucial ways that plants use to compensate for chemical energy synthesis.[3][4][5]
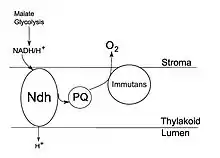
Chlororespiration – the latest model
Initially, the presence of chlororespiration as a legitimate respiratory process in plants was heavily doubted. However, experimentation on Chlamydomonas reinhardtii, discovered Plastoquinone (PQ) to be a redox carrier.[2] The role of this redox carrier is to transport electrons from the NAD(P)H enzyme to oxygen molecules on the thylakoid membrane.[6] Using this cyclic electron chain around photosystem one (PS I), chlororespiration compensates for the lack of light. This cyclic pathway also allows electrons to re-enter the PQ pool through NAD(P)H enzyme activity and production, which is then used to supply ATP molecules (energy) to plant cells.[7]
.jpg.webp)
In the year 2002, the discovery of the molecules; plastid terminal oxidase (PTOX) and NDH complexes have revolutionised the concept of chlororespiration.[2] Using evidence from experimentation on the plant species Rosa Meillandina, this latest model observes the role of PTOX to be an enzyme that prevents the PQ pool from over-reducing, by stimulating its reoxidation.[4] Whereas, the NDH complexes are responsible for providing a gateway for electrons to form an ETC.[4] The presence of such molecules are apparent in the non-appressed thylakoid membranes of higher order plants like Rosa Meillandina.[5][2][3]
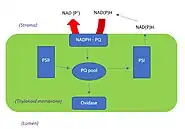
The relation between chlororespiration, photosynthesis and respiration
.jpg.webp)
Experimentation with respiratory oxidase inhibitors (for instance, cyanide) on unicellular algae has revealed interactive pathways to be present between chloroplasts and mitochondria. Metabolic pathways responsible for photosynthesis are present in chloroplasts, whereas respiratory metabolic pathways are present in mitochondria. In these pathways, metabolic carriers (like phosphate) exchange NAD(P)H molecules between photosynthetic and respiratory ETCs.[2] Evidence using mass spectrometry on algae and photosynthetic mutants of Chlamydomonas, discovered that oxygen molecules were also being exchanged between photosynthetic and chlororespiratory ETCs. [6] The mutant Chlamydomonas plant species, lacks photosystems one and two (PS I and PS II), so when the plant underwent flash-induced PS I activity, it resulted in no effect on mitochondrial pathways of respiration. Instead, this flash-induced PS I activity caused an exchange between photosynthetic and Chlororespiratory ETCs, which was observed using polarography.[6] This flash of PS I activity is triggered by an over-reduction of the PQ pool and/or lack of the pyridine nucleotide in the thylakoid membrane. A reduction in such molecules then stimulates NADPH and PTOX molecules to trigger chlororespiratory pathways.[6][2]
Furthermore, in the absence of light (and thus photosynthesis), chlororespiration plays an integral role in enabling metabolic pathways to compensate for chemical energy synthesis.[2] This is achieved through the oxidation of stromal compounds, which increases the PQ pool and allows for the chlororespiratory ETC to take place.[2][6]
Stimulation of chlororespiration
Quiles' experiment

An experiment on oat plants by scientist Maria Quiles, revealed that extreme light intensity can inhibit photosynthesis and result in the lack of PS II activity.[4] This reduction leads to an increase in NAD(P)H and PTOX levels which then causes the stimulation of chlororespiration.[4]
Oat leaves were incubated and Chlorophyll fluorescence emission was used to examine the effect of extreme light intensity.[4] As the emission of the Chlorophyll fluorescence increased the PQ pool decreased. This stimulated the cyclic electron flow, causing NAD(P)H and PTOX levels to ultimately incline and initiate the process of chlororespiration within the thylakoid membrane of oat plants.[4]
The effect of adding n-propyl gallate to the incubated leaves was also observed. N-propyl gallate is a molecule that helps distinguish between PQ reduction and oxidation activities by inhibiting PTOX.[8] Quiles noted an increase in chlorophyll fluorescence inside the thylakoid membrane of plant cells, after the addition of n-propyl gallate.[4] The result led to the stimulation of the NAD(P)H enzyme and its cyclic pathway; causing a continuous increase in chlorophyll fluorescence levels within the oat.[4]
Quiles' conclusion
After comparing the metabolic responses between oat plants under an average light intensity to that of oat plants under extreme light intensity, Quiles noted that the amount of PS II produced was of a lower amount in the leaves that underwent chlororespiration in extreme light.[4] Whereas higher levels of PS II were yielded by those leaves that underwent average light intensity. A higher of PS II is more efficient for chemical energy synthesis and thus for a plant's survival.[4] Quiles indicates that although the chlororespiratory pathway is less efficient, it still serves as a back-up response for energy production in plants.[4] Ultimately, Quiles concluded that the intense light on oat plants had caused PS II levels to reduce and thus, initiate an influx of gate-way (NAD(P)H) proteins to start the process of chlororespiration.[4]
Paredes' and Quiles' experiment
.jpg.webp)
Scientists Miriam Paredes and Maria Quiles led an investigation on the plant species Rosa Meillandina, and its metabolic response to water deficit. [3] They noted how limited water irrigation can cause a reduction in PS II levels, which then results in the inhibition of photosynthesis. Paredes and Quiles also noticed the increase in chlororespiration activity as a protective mechanism for the lack of photosynthesis.[3]
In the experiment, the plants in water deficit were analysed with fluorescence imaging technique. This form of analysis detected increased levels of PTOX, and NAD(P)H activity within the plant. [3] An increase in these two molecules led to the initiation of chlororespiration.[3]
N-propyl gallate was also added to these water deficit plants. The effect resulted in increased chlorophyll fluorescence levels. [3] Quiles recorded a similar outcome in the same species of plants that went under intense light. [4] This increase in chlorophyll fluorescence is attributed to the influx of NAD(P)H in the thylakoid membrane. [3] Which then led to an increase in the by-product, hydrogen peroxide, inside the thylakoid membrane.[3][4]
Paredes' and Quiles' conclusion
Paredes and Quiles concluded that chloroplasts under stress from water deficit rely on processes like the opening of stomata to disperse excess heat accumulated via metabolic processes within plant cells. [3] These metabolic processes are responsible for chemical energy synthesis that can be achieved via chlororespiratory ETCs when a reduction in photosynthesis activity is evident. [3]
Gasulla's, Casano's and Guéra's experiment
Scientists Francisco Gasulla, Leonardo Casano and Alfredo Guéra, observed the lichen plant's metabolic response when placed in dark conditions.[8] The light harvesting complex (LHC) inside the chloroplasts of Lichen is activated when subjected to darkness. [8] Gasulla, Casano and Guéra, noticed that this increase in LHC activity caused PS II and the PQ pool within lichen to decrease, indicating the initiation of chlororespiration.[8]
Immunodetection analysis was used to determine the amount of LHC molecules inside lichen in a dark environment, and a luminous environment. By determining the amount of LHC within the chloroplast, scientists were able to notice the reduction in PS II activity. This reduction was caused by a loss in excitation energy in the PS II ETC, which then stimulated an incline in chlororespiratory pathways. Gasulla, Casano and Guéra, gathered this result, when both light-adapted and dark-adapted lichen were placed in darkness. They found that the level of LHC molecules in dark-adapted lichen had doubled compared to light adapted lichen.[8] It was also noted that the chlororespiratory ETCs were triggered at a much earlier time in dark-adapted lichen, than compared to light-adapted lichen. [8]This resulted in a faster metabolic rate and chemical synthesis response in dark-adapted lichen due to chlororespiration.[8]
Gasulla's, Casano's and Guéra's conclusion
Gasulla, Casano and Guéra, concluded that the longer the lichen are subjected to darkness, the quicker the chlororespiratory pathways can begin.[8] This is due to the fast depletion of PTOX molecules which reduce the PQ pool. [8]These events then stimulate chlororespiratory ETCs into an ongoing loop until the lichen are placed in a luminous environment.[8]
They also derived LHC to be another indicator for chlororespiration. [8] When LHC concentrations inside the chloroplast increased, PS II activity decreased due to loss in ETC activity.[8] This reduction then stimulated chlororespiratory activity to compensate for chemical energy synthesis.[8]

Segura's and Quiles' experiment
An experiment observing chilling stress on the tropical plant species, Spathiphyllum wallisii by scientists Maria Segura and Maria Quiles, showcased varying responses by chlororespiratory pathways when different parts of the plant were chilled at 10 degrees Celsius.[9]
.JPG.webp)
Segura and Quiles noticed that when the roots of the plant were subjected to low temperatures (10 degrees Celsius), the level of chlororespiratory molecules (NADPH and PTOX) slightly varied when compared to the level of NADPH and PTOX within the controlled plant.[9] However, when the stem alone was cooled at 10 degrees Celsius, then the molecules NADPH, NDH and PTOX increased in amount as a result of reduced PS I activity.[9] Segura and Quiles then compared this result, by subjecting only the leaves of the plant to 10 degrees Celsius.[9] They noticed that this had caused the PS II activity to stop, and thus inhibit the process of photosynthesis.[9] The lack in photosynthetic activity, in combination with the incline in NADPH and PTOX molecules, then triggered chlororespiratory pathways to begin chemical energy synthesis.[9]
Furthermore, Segura and Quiles also noted that the simultaneous chilling of the leaves and heating of the roots (whilst the plant is under illumination), can cause the slowing and eventual inhibition of the ETC in PS II. [9] This then led to an over-reduction in the PQ pool., which ultimately stimulated chlororespiration.[9]
Segura and Quiles utilised the Fluorescence imaging technique to determine the level of photosynthetic activity in the leaves of the plants. By discerning the percentage of photosynthesis efficiency, Segura and Quiles were able to determine the likelihood of triggering chlororespiratory pathways. [9] They noticed that the percentage of photosynthesis efficiency remained high in test subjects where:
- only the leaves were chilled
- only the stem was chilled
- only the roots were chilled [9]
This high percentage of photosynthesis efficiency meant that the chances of chlororespiration taking place are slim. [9] However, this was not true for the plant that underwent both stem chilling at 10 degrees Celsius and root heating at 24 degrees Celsius.[9] The photosynthesis efficiency of this test subject was significantly lower when compared to the experimental control.[9] This also indicated the inhibition of PS II activity which then caused chlororespiration to begin.[9]
Segura and Quiles also used an immunoblot analysis to deduce the effect of varying temperatures on different parts of the plant. Specifically, the immunoblot measures the amount of PTOX and NDH complex accumulated within the thylakoid membrane of the chloroplast organelle.[9] An increase in NDH complex was evident in the plant where the stem was chilled at 10 degrees Celsius and the root heated at 24 degrees Celsius.[9] Chlororespiration was stimulated in this plant. [9] Dissimilarly, the immunoblot analysis detected no variation in the levels NDH complex and PTOX molecules in test subjects where:
- only the leaves were chilled
- only the stem was chilled
- only the roots were chilled[9]
These test subjects had similar concentrations of NDH and PTOX when compared to the concentration of NDH complex and PTOX molecules within the experimental control.[8][9]
Segura's and Quiles' conclusion
Segura and Quiles concluded that chilling stress only induces chlororespiration when the stem is significantly cool and the roots are simultaneously warmer compared to the average Spathiphyllum wallisii in controlled conditions.[9] Segura and Quiles notice that PS II is present in chloroplast, (which is lacking in roots), thus by chilling the stem (which contains chloroplast), PS II ETCs can then be inhibited to trigger a reduction in PQ pool and as a result, chlororespiration.[9]
Importance of chlororespiration
Although chlororespiration is not as efficient as photosynthesis in producing energy, [9] its significance its attributed to its role as a survival adaptation for plants when placed in conditions lacking light[8] and water[3] or if placed in uncomfortable temperatures[9][4] (note: optimum temperatures vary across different plant species).[9] Additionally, Cournac and Peltier noticed that chlororespiratory ETCs play a role in balancing electron flow across respiratory and photosynthetic ETCs.[2] This helps maintain water balance and regulate the plant's internal temperature.[2]
References
- Nixon, P. (2000). "Chlororespiration". Philosophical Transactions of the Royal Society B: Biological Sciences. 355 (1402): 355(1402), 1541–1547. doi:10.1098/rstb.2000.0714. PMC 1692878. PMID 11128007.
- Cournac, L.; Peltier, G. (2002). "Chlororespiration". Annual Review of Plant Biology. 53: 523–550.
- Paredes, Miriam; Quiles, María José (January 2013). "Stimulation of chlororespiration by drought under heat and high illumination in Rosa meillandina". Journal of Plant Physiology. 170 (2): 165–171. doi:10.1016/j.jplph.2012.09.010. PMID 23122789.
- Quiles, M. (2006). "Stimulation of chlororespiration by heat and high light intensity in oat plants". Plant, Cell & Environment. 29 (8): 1463–1470. doi:10.1111/j.1365-3040.2006.01510.x. PMID 16898010.
- Houille-Vernes, L.; Rappaport, F.; Wollman, F.-A.; Alric, J.; Johnson, X. (2011). "Plastid terminal oxidase 2 (PTOX2) is the major oxidase involved in chlororespiration in Chlamydomonas". Proceedings of the National Academy of Sciences of the United States of America. 108 (51): 20820–20825. Bibcode:2011PNAS..10820820H. doi:10.1073/pnas.1110518109. PMC 3251066. PMID 22143777.
- Peltier, G.; Schmidt, G. W. (1991). "Chlororespiration: an adaptation to nitrogen deficiency in Chlamydomonas reinhardtii". Proceedings of the National Academy of Sciences of the United States of America. 88 (11): 4791–4795. Bibcode:1991PNAS...88.4791P. doi:10.1073/pnas.88.11.4791. PMC 51752. PMID 11607187.
- Bennoun, P. (1982). "Evidence for a respiratory chain in the chloroplast". Proceedings of the National Academy of Sciences of the United States of America. 79 (14): 4352–4356. Bibcode:1982PNAS...79.4352B. doi:10.1073/pnas.79.14.4352. PMC 346669. PMID 16593210.
- Gasulla, Francisco; Casano, Leonardo; Guéra, Alfredo (2018). "Chlororespiration induces non‐photochemical quenching of chlorophyll fluorescence during darkness in lichen chlorobionts". Physiologia Plantarum. 166 (2): 538–552. doi:10.1111/ppl.12792. PMID 29952012.
- Segura, María V.; Quiles, María J. (March 2015). "Involvement of chlororespiration in chilling stress in the tropical species". Plant, Cell & Environment. 38 (3): 525–533. doi:10.1111/pce.12406. PMID 25041194.