LINGO1
Leucine rich repeat and Immunoglobin-like domain-containing protein 1[5] also known as LINGO-1 is a protein which is encoded by the LINGO1 gene in humans.[6][7] It belongs to the family of leucine-rich repeat proteins which are known for playing key roles[8] in the biology of the central nervous system. LINGO-1 is a functional component of the Nogo (neurite outgrowth inhibitor) receptor also known as the reticulon 4 receptor.




It has been suggested that LINGO-1 antagonists such as BIIB033[9] could significantly improve and regulate survival after neural injury caused by the protein.[10]
Structure

The human LINGO-1 is a single-pass type 1 transmembrane protein of 614 amino acids. It contains a signal sequence of 34 residues, followed by a LRR (leucine-rich repeat) domain, an Ig (immunoglobulin-like) domain, a stalk domain, a transmembrane region and a short cytoplasmic tail. As a transmembrane protein, it can mostly be found on the cell membrane.[11]
The LINGO-1 structure has been shown to be highly stable both in its crystal form and in solution, thanks to its leucine-rich repeat Ig-composite fold. Since the tetramer has a very large surface area into the cell membrane, it is thought that this may serve as an efficient and stable binding platform, facilitating the interaction with NgR, p75, TROY complex.
Extracellular domain
The extracellular domain consists of the signal sequence, 11 LRR motifs comprised between an N-terminal and C-terminal capping domains, and the Immunoglobulin-like (IgC2) domain.[7][12] The C-terminal LRR domain is essential for the protein's function with its screening for proteins that interact with this domain. The structure, together with biophysical analysis of LINGO-1 properties have revealed that the protein's LRR-Ig composite fold of the protein can drive it to associate with itself in a circular ring-like form, creating a closed and stable tetramer in solution and in crystal.
Intracellular domain
The intracellular part of the protein is formed by the transmembrane region and a cytoplasmic tail of 38 residues. It contains a canonical Epidermal Growth Factor Receptor (EGFR)-like tyrosine phosphorylation site on the 591 residue that is critical for intracellular signals.[13]
Co-receptor
LINGO-1 is a co-receptor that interacts with the ligand-binding Nogo-66 receptor (NogoR) in the Nogo receptor signaling complex.[12] The Nogo receptor complex is formed when Nogo-66 binds to its receptor.[14]
LINGO-1 is an homotetramer which forms a ternary complex with RTN4R/NGFR and RTN4R/TNFRSF19.
Post-translational modifications
LINGO-1 contains several N-glycosylation sites that could have a negative effect on its capacity to self-interact with cis or trans, with other partners, or gangliosides.[15] It also contains high-mannose glycans.
Tissue distribution
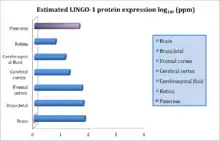
LINGO-1 is expressed almost exclusively in the central nervous system (CNS). It can be found in the brain and in neurons and oligodendrocytes. LINGO-1 mRNA is expressed in an almost exclusive manner in the central nervous system during both embryonic and postnatal stages. It is targeted to the plasma membrane of neurons, but it is possible that a smaller quantities of the protein may be found in other intracellular compartments.[16] Its highest expression is in specific adult human brain regions such as the cerebral cortex, a region involved in sensory-motor function, cognition and working memory; the hippocampus, responsible for long term memory and the encoding and retrieval of multi-sensory information; the amygdala, implicated in the stress response; as well as the thalamus, with a more constant and basal level of expression across the remainder of the brain.[17]
Function and mode of action
Since LINGO-1 is a leucine-rich repeat protein, which are known for their important role in protein-protein interactions in a wide variety of cellular processes and their implication in important functions like neuronal differentiation and growth or the regulation of axon guidance and regeneration processes, it is logical to deduce that its functions are linked with the nervous system.
LINGO-1 is an essential negative regulator of myelination. It has been implicated in the inhibition of axon regeneration through a ternary complex formed with NgR1/Nogo-66 (ligand-binding subunit) and p75 (signal transducing subunit). NgR1 relies on its co-receptors for transmembrane signalling. The three major myelin-associated inhibitory factors are Nogo, oligodendrocyte myelin glycoprotein, and myelin-associated glycoprotein which all share this trimolecular receptor complex. The inhibitory action is achieved through RhoA-GTP upregulation in response to the presence of MOG, MAG or Nogo-66 in the central nervous system.[12] LINGO-1 also inhibits oligodendrocyte precursor differentiation and myelination, by a mechanism that also involves activation of RhoA, but which apparently does not require p75 or NgR1.
LINGO-1 is involved in the regulation of neural apoptosis by inhibiting WNK3 kinase activity. It has been shown that blocking the extracellular domain of LINGO-1 disrupts the interaction between receptor kinases and LINGO-1 which directly attenuates inhibition of neuronal survival. However among the four WNK family members, only WNK3 has been shown to regulate and increase cell survival in a caspase-3-dependent pathway.[15][18]
To be able to understand how these components regulate signalling processes an experiment has been set up "model of serum deprivation" (SD) to prompt neuronal apoptosis. Research shows that treatments either with a construct containing the IgC2 or EGFR domains in the LINGO1 protein or with Nogo66 which act like a NgR1 agonist, therefore initiating a physiological response when combined with the receptor, resulting in an increased rate of apoptosis in primary cultured cortical neurons under SD.
In addition, reducing the expression levels of the serine/threonine Kinase WNK3 (using gene silencing via RNA interference (ShRNA)) or inhibiting its kinase activity had similar effects on the survival of such neurons. The adverse effects of Nogo66[19] have proved to enhance the co-association of LINGO1 and WNK3, causing the binding of WNK3 to the intracellular domain of LINGO1 leading to reduced WNK3 kinase activity. LINGO1 promotes neural apoptosis by inhibiting WNK3 kinase activity.[20]
Signaling pathways
LINGO-1 is able to interact with different co-factors and co-receptors, which can lead to the activation o signaling pathways that can have an effect on the regulation of neuronal survival, axon regeneration, oligodendrocyte differentiation, or myelination processes in the brain.[21]
Known interactions are with proteins such as Oligodendrocyte-myelinn glycoprotein, Nogo-A (neurotic outgrowth inhibitor), and myelin associated glycoproteins. LINGO-1 also interacts with transmembrane proteins: EFGR, along with its ligand epidermal growth factor (EFG); brain derived neurotrophic factor (BNDF) and its receptor, amyloid precursor protein (APP), and tropomyosin receptor kinase A (TrkA). There are other interactions with proteins that are implicated in neurological and psychiatric disorders: WNK lysine deficient protein in kinase 1 (WNK1), mitogen activated protein kinase 2/3 (MEK 2/3), extracellular signal reduced kinase 5 (ERK5), RhoA, and others.[22]
Neurological and psychiatric disorders
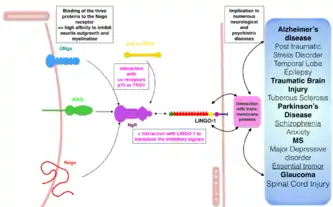
LINGO-1 is coded by the LINGO-1 gene, which is located on the human chromosome 15, more precisely on the locus 15q24-26, which is a region that has a primordial implication in number of psychiatric, addictive and anxiety related disorders. Genomic alterations of this regions can be factors for disorders such as schizophrenia, depression, autism, panic disorder or anxiety.[23]
Brain regions identified as highly expressing Lingo-1 transcripts have also been heavily implicated in both neurological and psychiatric disorders such as spinal cord injury, traumatic brain injury, multiple sclerosis (MS), Parkinson's disease, essential tremor (ET), Alzheimer's disease, epilepsy and glaucoma (central nervous system diseases); as well as stress and panic disorders, schizophrenia, amnesia, etc.[17] The role of Lingo-1 in these neurological disorders consists on its inhibitory role in neurite outgrowth, oligodendrocyte differentiation and myelination, making it difficult for the nervous system to regenerate the injured areas, whether these injuries come from endogenous or exogenous processes.
Spinal cord injury
Spinal cord injury results in the damage of the axonal tracts whose function is to control motor and sensory activity. This protein has been found in this axonal tracts of adolescent rat spinal cords following injury. Furthermore, a five time increase in Lingo-1 mRNA levels was detected 14 days post injury. Lingo-1-Fc, a soluble form of Lingo-1, has also been shown to antagonize Lingo-1 signaling pathways by inhibiting the binding of Lingo-1 to NgR, in consequence, vast improvements in the functional recovery of rats following lateral hemisection of the spinal cord were observed.[17]
Essential tremor and Parkinson
Essential tremor, one of the most common neurological diseases, is characterized by postural and action tremor. Recent research shows that around the 20% of people who suffer this disease have an increase of the protein LINGO1 in their cerebellum, therefore linking LINGO1 to essential tremor would result in the development of more effective symptomatic therapies and treatment.[24][25][26]
It has been found that there is a marker in LINGO-1 genome, a variant (rs9652490) that is significantly associated with essential tremor, increasing the risk of having the pathology.
As for Parkinson's disease, which is also an age-related movement disorder, it was discovers that levels of LINGO-1 are more elevated in the substantia nigra and cerebellum[27] of post-mortem Parkinson's disease brains compared to control groups. Dudem et al., (2020)[27] also demonstrated that LINGO1 is a novel regulatory subunit of large conductance, Ca2+ activated (BK) channels. It is thought that dopamine neuron survival and behavioral abnormalities are due to the over expression of LINGO-1 in Parkinson's patients.[17]
Traumatic brain injury
Traumatic brain injury involves the necrotic and apoptotic death of brain cells in vulnerable and delicate areas such as the cerebral cortex and hippocampus, where it is known that there is an expression of Lingo-1 in both development and the adult stage of life. RhoA signaling is largely responsible for the neuronal response to neuronal inhibitory proteins and the regeneration (or lack of in case of its activation) of damaged axons. Receptor Lingo-1 stimulates RhoA, which activates ROCK (RhoA kinase) which, in turn, stimulates LIM kinase, which then stimulates cofilin, which effectively reorganizes the actin cytoskeleton of the cell. In the case of neurons, activation of this pathway results in growth cone collapse, therefore inhibits the growth and repair of neural pathways and axons. Inhibition of this pathway by its various components usually results in some level of improved re-myelination.[28] The use of Lingo-1-Fc as an antagonist for Lingo-1 shows the inhibition of RhoA activation. Since this soluble form of Lingo-1 is able to block the interactions between Lingo-1 and NgR, it is reasonable to think that the blockade of RhoA occurs at the level of Lingo-1/NgR/p75 or TROY complex, leading to the conclusion that Lingo-1plays a very important part in the lack of re-myelination, repair of neural and axon injuries, etc.[17]
Schizophrenia
Schizophrenia is a chronic, severe and disabling brain disorder.
As said before, leucine-rich repeat and immunoglobulin domain-containing protein (Lingo-1) is an essential negative regulator of myelination and neurite extension. Both myelination and neurite outgrowth occur during brain maturation, and it is during this late period of brain development (adolescence and early adulthood) when schizophrenia is first expressed. In fact, myelination peaks during late adolescence, coinciding with the onset of schizophrenia. Consequently, an excessive action of Lingo-1 through demyelination and blocking neurite extension may be one of the possible causes of this disorder.
The brain regions which are highly disrupted in the pathophysiology of this disease are hippocampus and dorsolateral prefrontal cortex. Therefore, clinical studies have been developed in order to study these brain regions in people who suffer schizophrenia. To investigate the hypothesis that myelin fraction is lower in schizophrenia patients than in healthy subjects, a technique called magnetic resonance spectroscopy (MRS), which allows analysis of myelin, is used. This studies reported that there was in fact, a dysfunctional profile of myelination in these two areas of the brain in schizophrenia sufferers.[29]
Post-mortem studies were then realized in order to compare the levels of the protein Lingo-1 in these two brain regions (hippocampus and dorsolateral prefrontal cortex) between schizophrenia and healthy subjects. Effectively, it was shown that the levels of Lingo-1 where significantly higher in schizophrenia than in control groups.[30]
Taking this into account, there is a clear relationship in between schizophrenia and demyelination, therefore, this disease is linked with Lingo-1 protein. Very possibly, an effective treatment of this disease would be the use of Lingo-1 antagonists, such as Anti-Lingo-1, which would offset the lack of myelin and hopefully avoid the disease. Thus, this treatment is still in ways of development and research.[31]
Multiple sclerosis
Multiple sclerosis is among the most common neurological disorders in young adults and it consists in the destructions and damage of the central nervous system (CNS) myelin due to persistent inflammation in the brain and spinal cord. This demyelination is shown to cause mitochondrial dysfunction in axons, leading to their degeneration. These damages disrupt the ability and capacity of the CNS to communicate, causing, therefore, a wide range of symptoms including physical, mental and even psychiatric ones. The best way of re-myelination is encouraging the differentiation of endogenous adult precursor cells into mature oligodendrocytes in the injured regions. These precursor cells are called oligodendrocyte precursor cells (OPCs). It is known that in early stages of MS re-myelination can be achieved successfully and efficiently whereas it cannot in late and progressive stages. Regarding Lingo-1, we know that its signaling pathway is a negative regulator of OPCs differentiation, as well as Notch's and Wnt's.
Lingo-1 antagonists are able to promote re-myelination in CNS by means of stimulating OPCs differentiation which was before blocked by this protein. This has been seen in several experiments that resulted in significant increases of oligodendrocytes differentiation by targeting Lingo-1 with its antagonists, such as the antibody Anti-LINGO-1 (BIIB033).[32]
Glaucoma
Glaucoma is a group neurodegenerative diseases characterized by features including morphological changes in the optic nerve head and therefore in the visual fields of the patients. There are two main types; open-angle and closed-angle glaucoma. The loss of RGCs (retinal ganglion cells) and their axons results in visual field loss. Increasing evidence also supports the existence of compartmentalized degeneration in synapses. It has been shown that the first symptoms of this disease are usually ocular hypertension. Elevated IOP (intraocular pressure) has been identified as the etiology of glaucoma which causes neural RCG degeneration in the retina.[33]
LINGO1 was found to be expressed in the normal retina and was up regulated in RCGs after the induction of ocular hypertension in a rat chronic glaucoma model. Hence LINGO1 functions as a negative regulator of neuronal survival, axonal regeneration and oligodendrocyte differentiation. LINGO1 binds with TrkA and inhibits myelination by oligodendrocytes in vitro. Further more it binds to BDNF receptor and TrkB inhibiting the activation of TkrB by binding of BNDF after the induction of ocular hypertension.
Neuroprotection of RCGs
Even though BDNF is an important survival factor for RGCs both during development and adult life, BDNF can only slightly increase the survival rate of RCGs,[33] and does not significantly “rescue” injured RCGs in hypertensive eyes after episcleral vein cauterization. The negative regulatory function of LINGO1 may be involved in the limited neuroprotective effect of BDNF and it could be reversed after blocking LINGO1 function.
LINGO 1 negatively regulates TrkB activation through the signalling pathway of BDNF/TrKB, and anti-LINGO-1 exerts neuroprotective effects via activation of BDNF/TrkB.[34][35]
Better than BDNF and BII003 (LINGO1 antagonist) alone, the combined treatment of both provides long term RCG neuroprotection after the induction of ocular hypertension. In conclusion BII033 may provide an attractive therapeutic strategy to promote neuroprotection in glaucoma.[33]
Antagonists
Blocking the activity of lingo-1 has several potential applications in the treatment of neurodegenerative diseases.[22][36]
(A myelin sheath is a lipid protective coating that covers and protects nerve cells (axons). These sheaths makes possible rapid and accurate transmission of nerve signals. Multiple sclerosis destroys these myelin sheaths, leading to a deterioration in nerve signal transmission. Once this protective myelin coating is stripped away, it leads to apoptosis of the neuron; axons gradually die, causing the muscle spasms and paralysis that are characteristic of the disease.[37])
Anti-lingo-1 (BIIB033)
Anti-lingo-1 (BIIB033) is a monoclonal antibody specific to the lingo-1 protein and is designed to promote remyelination (the formation of new myelin on axons) and neuroprotection.[20][38] The protein lingo-1 inhibits the action of myelin-making cells, oligodendrocytes, which are surrounding the axons. Its antagonist, the antibody anti-lingo-1 would block this protein and even would be capable of myelin repair.
A number of clinical trials of the anti-lingo-1 antibody drug (BIIB033) have either been completed or are underway.[39] Acute optic neuritis (AON) is a disease which involves damage within the nerve fibers and loss of myelin within the optic nerve (it normally involves one eye and it's characterized by inflammation).[40] One clinical trial studying the effects on BIIB033 on acute optic neuritis.[41] Throughout the study, optic nerve conduction latency was measured (the time for a signal to travel from the retina to the brain's visual cortex).[42] As about half of patients with optic neuritis will later develop multiple sclerosis, BIIB033 antibody treatment is also being considered for the former disease. It is thought that Anti-Lingo-1 would produce the necessary myelin to avoid neurodegeneration.[9]
References
- GRCh38: Ensembl release 89: ENSG00000169783 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000049556 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "LINGO1 - Leucine-rich repeat and immunoglobulin-like domain-containing nogo receptor-interacting protein 1 - LINGO1 gene & protein". UniProt. Retrieved 2015-10-17.
- "LINGO1 leucine rich repeat and Ig domain containing 1". Entrez Gene.
- Carim-Todd L, Escarceller M, Estivill X, Sumoy L (Dec 2003). "LRRN6A/LERN1 (leucine-rich repeat neuronal protein 1) a novel gene with enriched expression in limbic system and neocortex". The European Journal of Neuroscience. 18 (12): 3167–82. doi:10.1111/j.1460-9568.2003.03003.x. PMID 14686891.
- de Wit J, Hong W, Luo L, Ghosh A (Jul 2011). "Role of leucine-rich repeat proteins in the development and function of neural circuits" (PDF). Annual Review of Cell and Developmental Biology. 27: 697–729. doi:10.1146/annurev-cellbio-092910-154111. PMID 21740233.
- "BIIB033". Multiple Sclerosis Discovery Forum. Retrieved 2015-10-18.
- Agúndez JA, Jiménez-Jimenez FJ, Alonso-Navarro H, García-Martín E (2015). "The potential of LINGO-1 as a therapeutic target for essential tremor". Expert Opin. Ther. Targets. 19 (8): 1139–48. doi:10.1517/14728222.2015.1028360. PMID 25862159.
- Mosyak L, Wood A, Dwyer B, Buddha M, Johnson M, Aulabaugh A, Zhong X, Presman E, Benard S, Kelleher K, Wilhelm J, Stahl ML, Kriz R, Gao Y, Cao Z, Ling HP, Pangalos MN, Walsh FS, Somers WS (Nov 2006). "The structure of the Lingo-1 ectodomain, a module implicated in central nervous system repair inhibition". The Journal of Biological Chemistry. 281 (47): 36378–90. doi:10.1074/jbc.M607314200. PMID 17005555.
- Mi S, Lee X, Shao Z, Thill G, Ji B, Relton J, Levesque M, Allaire N, Perrin S, Sands B, Crowell T, Cate RL, McCoy JM, Pepinsky RB (Mar 2004). "LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex". Nature Neuroscience. 7 (3): 221–8. doi:10.1038/nn1188. PMID 14966521.
- Llorens F, Gil V, Iraola S, Carim-Todd L, Martí E, Estivill X, Soriano E, del Rio JA, Sumoy L (2008). "Developmental analysis of Lingo-1/Lern1 protein expression in the mouse brain: interaction of its intracellular domain with Myt1l". Developmental Neurobiology. 68 (4): 521–41. doi:10.1002/dneu.20607. PMID 18186492.
- Alhoshani A, Vithayathil R, Weiss GA, Cocco MJ (Jan 2014). "Model of the Nogo: Nogo Receptor Complex". Biophysical Journal. 106 (22, Supplement 1): 410a–411a. doi:10.1016/j.bpj.2013.11.2311.
- Jepson S, Vought B, Gross CH, Gan L, Austen D, Frantz JD, Zwahlen J, Lowe D, Markland W, Krauss R (Jun 2012). "LINGO-1, a transmembrane signaling protein, inhibits oligodendrocyte differentiation and myelination through intercellular self-interactions". The Journal of Biological Chemistry. 287 (26): 22184–95. doi:10.1074/jbc.M112.366179. PMC 3381180. PMID 22514275.
- "LINGO1 Gene". Gene Cards. Retrieved 2015-10-18.
- Andrews JL, Fernandez-Enright F (2015). "A decade from discovery to therapy: Lingo-1, the dark horse in neurological and psychiatric disorders". Neuroscience & Biobehavioral Reviews. 56: 97–114. doi:10.1016/j.neubiorev.2015.06.009. PMID 26143511.
- Veríssimo F, Silva E, Morris JD, Pepperkok R, Jordan P (Jul 2006). "Protein kinase WNK3 increases cell survival in a caspase-3-dependent pathway". Oncogene. 25 (30): 4172–82. doi:10.1038/sj.onc.1209449. PMID 16501604.
- Lee H, Raiker SJ, Venkatesh K, Geary R, Robak LA, Zhang Y, Yeh HH, Shrager P, Giger RJ (Mar 2008). "Synaptic function for the Nogo-66 receptor NgR1: regulation of dendritic spine morphology and activity-dependent synaptic strength". The Journal of Neuroscience. 28 (11): 2753–65. doi:10.1523/JNEUROSCI.5586-07.2008. PMC 6670664. PMID 18337405.
- Zhang Z, Xu X, Xiang Z, Yu Z, Feng J, He C (Apr 2013). "LINGO-1 receptor promotes neuronal apoptosis by inhibiting WNK3 kinase activity". The Journal of Biological Chemistry. 288 (17): 12152–60. doi:10.1074/jbc.M112.447771. PMC 3636899. PMID 23482566.
- "Lingo1 protein (Mus musculus) - STRING network view". string-db.org. Retrieved 2015-10-30.
- Andrews JL, Fernandez-Enright F (2015). "A decade from discovery to therapy: Lingo-1, the dark horse in neurological and psychiatric disorders". Neurosci Biobehav Rev. 56: 97–114. doi:10.1016/j.neubiorev.2015.06.009. PMID 26143511.
- "Chromosome 15". Genetics Home Reference. 2015-10-26. Retrieved 2015-10-30.
- Delay C, Tremblay C, Brochu E, Paris-Robidas S, Emond V, Rajput AH, Rajput A, Calon F (Nov 2014). "Increased LINGO1 in the cerebellum of essential tremor patients". Movement Disorders. 29 (13): 1637–47. doi:10.1002/mds.25819. PMID 24531928.
- Kuo SH, Tang G, Louis ED, Ma K, Babji R, Balatbat M, Cortes E, Vonsattel JP, Yamamoto A, Sulzer D, Faust PL (Jun 2013). "Lingo-1 expression is increased in essential tremor cerebellum and is present in the basket cell pinceau". Acta Neuropathologica. 125 (6): 879–89. doi:10.1007/s00401-013-1108-7. PMC 3663903. PMID 23543187.
- Tan EK (2010). "LINGO1 and essential tremor: linking the shakes. Linking LINGO1 to essential tremor". Eur. J. Hum. Genet. 18 (7): 739–40. doi:10.1038/ejhg.2010.25. PMC 2987372. PMID 20372187.
- Dudem S, Large RJ, Kulkarni S, McClafferty H, Tikhonova IG, Sergeant GP, et al. (January 2020). "2+-activated potassium channels". Proceedings of the National Academy of Sciences of the United States of America. 117 (4): 2194–2200. doi:10.1073/pnas.1916715117. PMID 31932443.
- Yiu G, He Z (Aug 2006). "Glial inhibition of CNS axon regeneration". Nature Reviews. Neuroscience. 7 (8): 617–27. doi:10.1038/nrn1956. PMC 2693386. PMID 16858390.
- "Schizophrenia". www.nimh.nih.gov. The National Institute of Mental Health (NIMH), U.S. Department of Health and Human Services. Retrieved 2015-10-28.
- Flynn SW, Lang DJ, Mackay AL, Goghari V, Vavasour IM, Whittall KP, Smith GN, Arango V, Mann JJ, Dwork AJ, Falkai P, Honer WG (Sep 2003). "Abnormalities of myelination in schizophrenia detected in vivo with MRI, and post-mortem with analysis of oligodendrocyte proteins". Molecular Psychiatry. 8 (9): 811–20. doi:10.1038/sj.mp.4001337. PMID 12931208.
- Fernandez-Enright F, Andrews JL, Newell KA, Pantelis C, Huang XF (2014-01-21). "Novel implications of Lingo-1 and its signaling partners in schizophrenia". Translational Psychiatry. 4 (1): e348. doi:10.1038/tp.2013.121. PMC 3905231. PMID 24448210.
- Robson J, Trounson A (Sep 2011). "The potential of stem cells for 21st century neuroscience". Neurochemistry International. 59 (3): 327–8. doi:10.1016/J.NEUINT.2011.07.011. PMID 21827925.
- Fu Q, So K (2015). "Chapter 17 – Neuroprotection of Retinal Ganglion Cells in Glaucoma by Blocking LINGO-1 Function or Using a Nogo-66 Receptor Antagonist". In So K, Xu X (eds.). Neural regeneration. USA: Elsevier Inc. pp. 263–271. doi:10.1016/B978-0-12-801732-6.00017-3. ISBN 978-0-12-801732-6.
- Naylor RL, Robertson AG, Allen SJ, Sessions RB, Clarke AR, Mason GG, Burston JJ, Tyler SJ, Wilcock GK, Dawbarn D (Mar 2002). "A discrete domain of the human TrkB receptor defines the binding sites for BDNF and NT-4". Biochemical and Biophysical Research Communications. 291 (3): 501–7. doi:10.1006/bbrc.2002.6468. PMID 11855816.
- Haniu M, Montestruque S, Bures EJ, Talvenheimo J, Toso R, Lewis-Sandy S, Welcher AA, Rohde MF (Oct 1997). "Interactions between brain-derived neurotrophic factor and the TRKB receptor. Identification of two ligand binding domains in soluble TRKB by affinity separation and chemical cross-linking". The Journal of Biological Chemistry. 272 (40): 25296–303. doi:10.1074/jbc.272.40.25296. PMID 9312147.
- Mi S, Pepinsky RB, Cadavid D (2013). "Blocking LINGO-1 as a therapy to promote CNS repair: from concept to the clinic". CNS Drugs. 27 (7): 493–503. doi:10.1007/s40263-013-0068-8. PMID 23681979.
- Herper M (2015-10-25). "Biogen MS Drug Results Are 'Mildly Encouraging' In Eye Disease Test". Forbes. Retrieved 2015-10-29.
- Deiß A, Brecht I, Haarmann A, Buttmann M (2013). "Treating multiple sclerosis with monoclonal antibodies: a 2013 update". Expert Review of Neurotherapeutics. 13 (3): 313–35. doi:10.1586/ern.13.17. PMID 23448220.
- "BIIB033 Clinical Trials". ClinicalTrials.gov.
- Shams PN, Plant GT (Sep 2009). "Optic neuritis: a review". International MS Journal. 16 (3): 82–9. PMID 19878630.
- Clinical trial number NCT01721161 for "BIIB033 In Acute Optic Neuritis (AON) (RENEW)" at ClinicalTrials.gov
- "Anti-LINGO-1". Multiple Sclerosis Society. Archived from the original on 2015-12-22. Retrieved 2015-10-29.
Further reading
- Saha N, Kolev M, Nikolov DB (2014). "Structural features of the Nogo receptor signaling complexes at the neuron/myelin interface". Neuroscience Research. 87: 1–7. doi:10.1016/j.neures.2014.06.003. PMID 24956133.
- Carim-Todd L, Escarceller M, Estivill X, Sumoy L (Dec 2003). "LRRN6A/LERN1 (leucine-rich repeat neuronal protein 1), a novel gene with enriched expression in limbic system and neocortex". The European Journal of Neuroscience. 18 (12): 3167–82. doi:10.1111/j.1460-9568.2003.03003.x. PMID 14686891.
- Mi S, Lee X, Shao Z, Thill G, Ji B, Relton J, Levesque M, Allaire N, Perrin S, Sands B, Crowell T, Cate RL, McCoy JM, Pepinsky RB (Mar 2004). "LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex". Nature Neuroscience. 7 (3): 221–8. doi:10.1038/nn1188. PMID 14966521.
- Shao Z, Browning JL, Lee X, Scott ML, Shulga-Morskaya S, Allaire N, Thill G, Levesque M, Sah D, McCoy JM, Murray B, Jung V, Pepinsky RB, Mi S (Feb 2005). "TAJ/TROY, an orphan TNF receptor family member, binds Nogo-66 receptor 1 and regulates axonal regeneration". Neuron. 45 (3): 353–9. doi:10.1016/j.neuron.2004.12.050. PMID 15694322.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (Oct 2005). "Towards a proteome-scale map of the human protein-protein interaction network". Nature. 437 (7062): 1173–8. doi:10.1038/nature04209. PMID 16189514.
- Mosyak L, Wood A, Dwyer B, Buddha M, Johnson M, Aulabaugh A, Zhong X, Presman E, Benard S, Kelleher K, Wilhelm J, Stahl ML, Kriz R, Gao Y, Cao Z, Ling HP, Pangalos MN, Walsh FS, Somers WS (Nov 2006). "The structure of the Lingo-1 ectodomain, a module implicated in central nervous system repair inhibition". The Journal of Biological Chemistry. 281 (47): 36378–90. doi:10.1074/jbc.M607314200. PMID 17005555.
- Satoh J, Tabunoki H, Yamamura T, Arima K, Konno H (Feb 2007). "TROY and LINGO-1 expression in astrocytes and macrophages/microglia in multiple sclerosis lesions". Neuropathology and Applied Neurobiology. 33 (1): 99–107. doi:10.1111/j.1365-2990.2006.00787.x. PMID 17239012.
- Inoue H, Lin L, Lee X, Shao Z, Mendes S, Snodgrass-Belt P, Sweigard H, Engber T, Pepinsky B, Yang L, Beal MF, Mi S, Isacson O (Sep 2007). "Inhibition of the leucine-rich repeat protein LINGO-1 enhances survival, structure, and function of dopaminergic neurons in Parkinson's disease models". Proceedings of the National Academy of Sciences of the United States of America. 104 (36): 14430–5. doi:10.1073/pnas.0700901104. PMC 1955463. PMID 17726113.
External links
- Overview of all the structural information available in the PDB for UniProt: Q96FE5 (Leucine-rich repeat and immunoglobulin-like domain-containing nogo receptor-interacting protein 1) at the PDBe-KB.
PDB gallery | |
|---|---|
|

