Membrane vesicle trafficking
Membrane vesicle trafficking in eukaryotic animal cells involves movement of important biochemical signal molecules from synthesis-and-packaging locations in the Golgi body to specific 'release' locations on the inside of the plasma membrane of the secretory cell, in the form of Golgi membrane-bound micro-sized vesicles, termed membrane vesicles (MVs). In this process, the 'packed' cellular products are released/secreted outside the cell across its plasma membrane. However, this vesicular membrane is retained and recycled by the secretory cells. This phenomenon has a key role in synaptic neurotransmission, endocrine secretion, mucous secretion, granular-product secretion by neutrophils, etc. The scientists behind this discovery were awarded Nobel prize for the year 2013. In the prokaryotic gram-negative bacterial cells, membrane vesicle trafficking is mediated via bacterial outer membrane bounded nano-sized vesicles, called bacterial outer membrane vesicles (OMVs). In this case, however, the OMV membrane is secreted as well, along with OMV-contents to outside the secretion-active bacterium. This phenomenon has a key role in host-pathogen interactions, endotoxic shock in patients, invasion and infection of animals/plants, inter-species bacterial competition, quorum sensing, exocytosis, etc.
Movement within eukaryotic cells
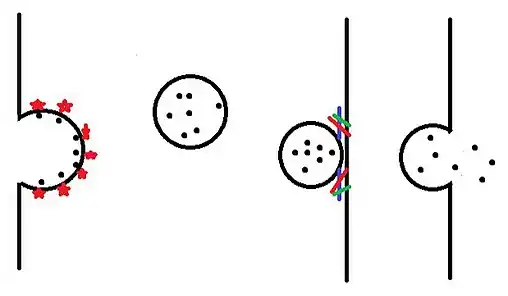
Once vesicles are produced in the endoplasmic reticulum and modified in the golgi body they make their way to a variety of destinations within the cell. Vesicles first leave the golgi body and are released into the cytoplasm in a process called budding. Vesicles are then moved towards their destination by motor proteins. Once the vesicle arrives at its destination it joins with the bi-lipid layer in a process called fusion, and then releases its contents.
Budding
Receptors embedded in the membrane of the golgi body bind specific cargo (such as dopamine) on the lumenal side of the vesicle. These cargo receptors then recruit a variety of proteins including other cargo receptors and coat proteins such as clathrin, COPI and COPII. As more and more of these coating proteins come together, they cause the vesicle to bud outward and eventually break free into the cytoplasm. The coating proteins are then shed into the cytoplasm to be recycled and reused.[1]
Motility between cell compartments
For movement between different compartments within the cell, vesicles rely on the motor proteins myosin, kinesin (primarily anterograde transport) and dynein (primarily retrograde transport). One end of the motor proteins attaches to the vesicle while the other end attaches to either microtubules or microfilaments. The motor proteins then move by hydrolyzing ATP, which propels the vesicle towards its destination.[2]
Docking and Fusion
As a vesicle nears its intended location, RAB proteins in the vesicle membrane interact with docking proteins at the destination site. These docking proteins bring the vesicle in closer to interact with the SNARE Complex found in the target membrane. The SNARE complex reacts with synaptobrevin found on the vesicle membrane.[3] This forces the vesicle membrane against the membrane of the target complex (or the outer membrane of the cell) and causes the two membranes to fuse. Depending on whether the vesicle fuses with a target complex or the outer membrane, the contents of the vesicle are then released either into the target complex or outside the cell.[4]
Examples In eukaryotes
- Intracellular trafficking occurs between subcellular compartments like Golgi cisternae and multivesicular endosomes for transport of soluble proteins as MVs.
- Budding of MVs directly from plasma membrane as microvesicles released outside the secretory cells.
- Exosomes are MVs that can form inside an internal compartment like multivesicular endosome. Exosomes are released eventually due to fusion of this endosome with plasma membrane of cell.
- Hijacking of exosomal machinery by some viruses like retroviruses, wherein viruses bud inside multivesicular endosomes and get secreted subsequently as exosomes.
All these types (1-4) of modes of membrane vesicle trafficking, taking place in eukaryotic cells have been explained diagrammatically.[5]
In prokaryotes
Unlike in eukaryotes, membrane vesicular trafficking in prokaryotes is an emerging area in interactive biology for intra-species (quorum sensing) and inter-species signaling at host-pathogen interface, as prokaryotes lack internal membrane-compartmentalization of their cytoplasm.
For more than four decades, cultures of gram negative microbes revealed the presence of nanoscale membrane vesicles. A role for membrane vesicles in pathogenic processes has been suspected since the 1970s, when they were observed in gingival plaque by electron microscopy.[6] These vesicles were suspected to promote bacterial adhesion to the host epithelial cell surface.[7] Their role in invasion of animal host cells in vivo was then demonstrated.[8] In inter-bacterial interactions, OMVs released by Pseudomonas aeruginosa microbes were shown to fuse with outer membrane of other gram negative microbes causing their bacteriolysis; these OMVs could lyse gram-positive microbes as well.[9] Role of OMVs in Helicobacter pylori infection of human primary antral epithelial cells, as model that closely resembles human stomach, has also been confirmed[10] VacA-containing OMVs could also be detected in human gastric mucosa, infected with H. pylori..[11] Salmonella OMVs were also shown to have direct role in invasion of chicken ileal epithelial cells in vivo in the year, 1993 (ref 4) and later, in hijacking of defense macrophages into sub-service for pathogen replication and consequent apoptosis of infected macrophages in typhoid-like animal infection.[12] These studies brought the focus on OMVs into membrane vesicle trafficking and showed this phenomenon as involved in multifarious processes like genetic transformation, quorum sensing, competition arsenal among microbes, etc., and invasion, infection, immuno-modulation, etc., of animal hosts.[6] A mechanism has already been proposed for generation of OMVs by gram negative microbes involving, expansion of pockets of periplasm (named, periplasmic organelles) due to accumulation of bacterial cell secretions and their pinching off as outer membrane bounded vesicles (OMVs) on the lines of a 'soap bubble' formation with a bubble tube, and further fusion or uptake of diffusing OMVs by host/target cells (Fig. 2).[13]
.png.webp)
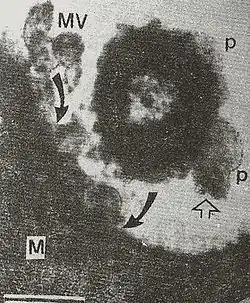
In conclusion, membrane vesicle trafficking via OMVs of Gram-negative organisms, cuts across species and kingdoms - including plant kingdom[14] - in the realm of cell-to-cell signaling.
See also
References
- Bonifacino, Juan (January 2004). "The Mechanisms of Vesicle Budding and Fusion". Cell. 116 (2): 153–166. doi:10.1016/S0092-8674(03)01079-1. PMID 14744428.
- Hehnly H, Stamnes M (May 2007). "Regulating cytoskeleton-based vesicle motility". FEBS Letters. 581 (11): 2112–8. doi:10.1016/j.febslet.2007.01.094. PMC 1974873. PMID 17335816.
- Nanavati C, Markin VS, Oberhauser AF, Fernandez JM (October 1992). "The exocytotic fusion pore modeled as a lipidic pore". Biophysical Journal. 63 (4): 1118–32. Bibcode:1992BpJ....63.1118N. doi:10.1016/S0006-3495(92)81679-X. PMC 1262250. PMID 1420930.
- Papahadjopoulos D, Nir S, Düzgünes N (April 1990). "Molecular mechanisms of calcium-induced membrane fusion". Journal of Bioenergetics and Biomembranes. 22 (2): 157–79. doi:10.1007/BF00762944. PMID 2139437.
- Théry C, Ostrowski M, Segura E (August 2009). "Membrane vesicles as conveyors of immune responses". Nature Reviews. Immunology. 9 (8): 581–93. doi:10.1038/nri2567. PMID 19498381.
- Ellis TN, Kuehn MJ (March 2010). "Virulence and immunomodulatory roles of bacterial outer membrane vesicles". Microbiology and Molecular Biology Reviews. 74 (1): 81–94. doi:10.1128/MMBR.00031-09. PMC 2832350. PMID 20197500.
- Halhoul N, Colvin JR (February 1975). "The ultrastructure of bacterial plaque attached to the gingiva of man". Archives of Oral Biology. 20 (2): 115–8. doi:10.1016/0003-9969(75)90164-8. PMID 1054578.
- YashRoy RC (1993). "Electron microscope studies of surface pili and vesicles of Salmonella 3,10:r:- organisms". Indian Journal of Animal Sciences. 63 (2): 99–102.
- Kadurugamuwa JL, Beveridge TJ (May 1996). "Bacteriolytic effect of membrane vesicles from Pseudomonas aeruginosa on other bacteria including pathogens: conceptually new antibiotics". Journal of Bacteriology. 178 (10): 2767–74. doi:10.1128/jb.178.10.2767-2774.1996. PMC 178010. PMID 8631663.
- Heczko U, Smith VC, Mark Meloche R, Buchan AM, Finlay BB (November 2000). "Characteristics of Helicobacter pylori attachment to human primary antral epithelial cells". Microbes and Infection. 2 (14): 1669–76. doi:10.1016/s1286-4579(00)01322-8. PMID 11137040.
- Fiocca R, Necchi V, Sommi P, Ricci V, Telford J, Cover TL, Solcia E (June 1999). "Release of Helicobacter pylori vacuolating cytotoxin by both a specific secretion pathway and budding of outer membrane vesicles. Uptake of released toxin and vesicles by gastric epithelium". The Journal of Pathology. 188 (2): 220–6. doi:10.1002/(sici)1096-9896(199906)188:2<220::aid-path307>3.0.co;2-c. PMID 10398168.
- Yashroy RC (2000). "Hijacking of macrophages by Salmonella (3,10:r:-) through 'type-III' secretion-like exocytotic signaling: a mechanism for infection of chicken ileum". Indian Journal of Poultry Science. 35 (3): 276–281.
- YashRoy RC (June 2003). "Eucaryotic cell intoxication by gram-negative pathogens: a novel bacterial outermembrane-bound nanovesicular exocytosis model for type-III secretion system". Toxicology International. 10 (1): 1–9.
- Bahar O, Pruitt R, Luu DD, Schwessinger B, Daudi A, Liu F, Ruan R, Fontaine-Bodin L, Koebnik R, Ronald P (2014). "The Xanthomonas Ax21 protein is processed by the general secretory system and is secreted in association with outer membrane vesicles". PeerJ. 2: e242. doi:10.7717/peerj.242. PMC 3897388. PMID 24482761.
External links
- Nobel Prize of year 2013 in Physiology and Medicine - press release http://www.nobelprize.org/nobel_prizes/medicine/laureates/2013/press.html
- Discovery of vesicular exocytosis in prokaryotes https://www.researchgate.net/publication/230793568_Discovery_of_vesicular_exocytosis_in_prokaryotes_and_its_role_in_Salmonella_invasion?ev=prf_pub