Cell adhesion
Cell adhesion is the process by which cells interact and attach to neighbouring cells through specialised molecules of the cell surface. This process can occur either through direct contact between cell surfaces such as cell junctions or indirect interaction, where cells attach to surrounding extracellular matrix, a gel-like structure containing molecules released by cells into spaces between them.[1] Cells adhesion occurs from the interactions between cell-adhesion molecules (CAMs),[2] transmembrane proteins located on the cell surface. Cell adhesion links cells in different ways and can be involved in signal transduction for cells to detect and respond to changes in the surroundings.[1][3] Other cellular processes regulated by cell adhesion include cell migration and tissue development in multicellular organisms.[4] Alterations in cell adhesion can disrupt important cellular processes and lead to a variety of diseases, including cancer[5][6] and arthritis.[7] Cell adhesion is also essential for infectious organisms, such as bacteria or viruses, to cause diseases.[8][9]
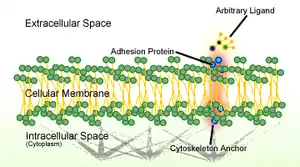
General mechanism

CAMs are classified into four major families: integrins, immunoglobulin (Ig) superfamily, cadherins, and selectins.[2] Cadherins and IgSF are homophilic CAMs, as they directly bind to the same type of CAMs on another cell, while integrins and selectins are heterophilic CAMs that bind to different types of CAMs.[2] Each of these adhesion molecules has a different function and recognizes different ligands. Defects in cell adhesion are usually attributable to defects in expression of CAMs.
In multicellular organisms, bindings between CAMs allow cells to adhere to one another and creates structures called cell junctions. According to their functions, the cell junctions can be classified as:[1]
- Anchoring junctions (adherens junctions, desmosomes and hemidesmosomes), which maintain cells together and strengthens contact between cells.
- Occluding junctions (tight junctions), which seal gaps between cells through cell–cell contact, making an impermeable barrier for diffusion
- Channel-forming junctions (gap junctions), which links cytoplasm of adjacent cells allowing transport of molecules to occur between cells
- Signal-relaying junctions, which can be synapses in the nervous system
Alternatively, cell junctions can be categorised into two main types according to what interacts with the cell: cell–cell junctions, mainly mediated by cadherins, and cell–matrix junctions, mainly mediated by integrins.
Cell–cell junctions
Cell–cell junctions can occur in different forms. In anchoring junctions between cells such as adherens junctions and desmosomes, the main CAMs present are the cadherins. This family of CAMs are membrane proteins that mediate cell–cell adhesion through its extracellular domains and require extracellular Ca2+ ions to function correctly.[2] Cadherins forms homophilic attachment between themselves, which results in cells of a similar type sticking together and can lead to selective cell adhesion, allowing vertebrate cells to assemble into organised tissues.[1] Cadherins are essential for cell–cell adhesion and cell signalling in multicellular animals and can be separated into two types: classical cadherins and non-classical cadherins.[2]
Adherens junctions
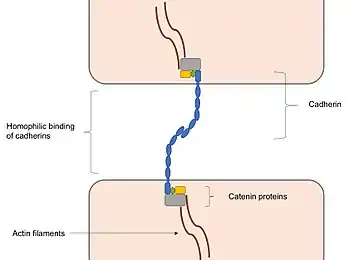
Adherens junctions mainly function to maintain the shape of tissues and to hold cells together. In adherens junctions, cadherins between neighbouring cells interact through their extracellular domains, which share a conserved calcium-sensitive region in their extracellular domains. When this region comes into contact with Ca2+ ions, extracellular domains of cadherins undergo a conformational change from the inactive flexible conformation to a more rigid conformation in order to undergo homophilic binding. Intracellular domains of cadherins are also highly conserved, as they bind to proteins called catenins, forming catenin-cadherin complexes. These protein complexes link cadherins to actin filaments. This association with actin filaments is essential for adherens junctions to stabilise cell–cell adhesion.[10][11][12] Interactions with actin filaments can also promote clustering of cadherins, which are involved in the assembly of adherens junctions. This is since cadherin clusters promote actin filament polymerisation ,which in turn promotes the assembly of adherens junctions by binding to the cadherin–catenin complexes that then form at the junction.
Desmosomes
Desmosomes are structurally similar to adherens junctions but composed of different components. Instead of classical cadherins, non-classical cadherins such as desmogleins and desmocollins act as adhesion molecules and they are linked to intermediate filaments instead of actin filaments.[13] No catenin is present in desmosomes as intracellular domains of desmosomal cadherins interact with desmosomal plaque proteins, which form the thick cytoplasmic plaques in desmosomes and link cadherins to intermediate filaments.[14] Desmosomes provides strength and resistance to mechanical stress by unloading forces onto the flexible but resilient intermediate filaments, something that cannot occur with the rigid actin filaments.[13] This makes desmosomes important in tissues that encounter high levels of mechanical stress, such as heart muscle and epithelia, and explains why it appears frequently in these types of tissues.
Tight junctions
Tight junctions are normally present in epithelial and endothelial tissues, where they seal gaps and regulate paracellular transport of solutes and extracellular fluids in these tissues that function as barriers.[15] Tight junction is formed by transmembrane proteins, including claudins, occludins and tricellulins, that bind closely to each other on adjacent membranes in a homophilic manner.[1] Similar to anchoring junctions, intracellular domains of these tight junction proteins are bound with scaffold proteins that keep these proteins in clusters and link them to actin filaments in order to maintain structure of the tight junction.[16] Claudins, essential for formation of tight junctions, form paracellular pores which allow selective passage of specific ions across tight junctions making the barrier selectively permeable.[15]
Gap junctions
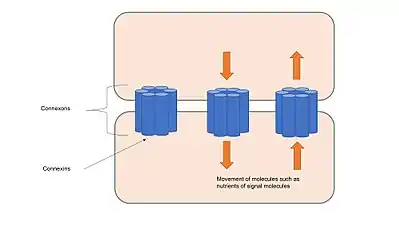
Gap junctions are composed of channels called connexons, which consist of transmembrane proteins called connexins clustered in groups of six.[17] Connexons from adjacent cells form continuous channels when they come into contact and align with each other. These channels allow transport of ions and small molecules between cytoplasm of two adjacent cells, apart from holding cells together and provide structural stability like anchoring junctions or tight junctions.[1] Gap junction channels are selectively permeable to specific ions depending on which connexins form the connexons, which allows gap junctions to be involved in cell signalling by regulating the transfer of molecules involved in signalling cascades.[18] Channels can respond to many different stimuli and are regulated dynamically either by rapid mechanisms, such as voltage gating, or by slow mechanism, such as altering numbers of channels present in gap junctions.[17]
Adhesion mediated by selectins
Selectins are a family of specialised CAMs involved in transient cell–cell adhesion occurring in the circulatory system. They mainly mediate the movement of white blood cells (leukocytes) in the bloodstream by allowing the white blood cells to "roll" on endothelial cells through reversible bindings of selections.[19] Selectins undergo heterophilic bindings, as its extracellular domain binds to carbohydrates on adjacent cells instead of other selectins, while it also require Ca2+ ions to function, same as cadherins.[1] cell–cell adhesion of leukocytes to endothelial cells is important for immune responses as leukocytes can travel to sites of infection or injury through this mechanism.[20] At these sites, integrins on the rolling white blood cells are activated and bind firmly to the local endothelial cells, allowing the leukocytes to stop migrating and move across the endothelial barrier.[20]
Adhesion mediated by members of the immunoglobulin superfamily
The immunoglobulin superfamily (IgSF) is one of the largest superfamily of proteins in the body and it contains many diverse CAMs involved in different functions. These transmembrane proteins have one or more immunoglobulin-like domains in their extracellular domains and undergo calcium-independent binding with ligands on adjacent cells.[21] Some IgSF CAMs, such as neural cell adhesion molecules (NCAMs), can perform homophilic binding while others, such as intercellular cell adhesion molecules (ICAMs) or vascular cell adhesion molecules (VCAMs) undergo heterophilic binding with molecules like carbohydrates or integrins.[22] Both ICAMs and VCAMs are expressed on vascular endothelial cells and they interact with integrins on the leukocytes to assist leukocyte attachment and its movement across the endothelial barrier.[22]
Cell–matrix junctions
Cells creates extracellular matrix by releasing molecules into its surrounding extracellular space. Cells have specific CAMs that will bind to molecules in the extracellular matrix and link the matrix to the intracellular cytoskeleton.[1] Extracellular matrix can act as a support when organising cells into tissues and can also be involved in cell signalling by activating intracellular pathways when bound to the CAMs.[2] Cell–matrix junctions are mainly mediated by integrins, which also clusters like cadherins to form firm adhesions. Integrins are transmembrane heterodimers formed by different α and β subunits, both subunits with different domain structures.[23] Integrins can signal in both directions: inside-out signalling, intracellular signals modifying the intracellular domains, can regulate affinity of integrins for their ligands, while outside-in signalling, extracellular ligands binding to extracellular domains, can induce conformational changes in integrins and initiate signalling cascades.[23] Extracellular domains of integrins can bind to different ligands through heterophilic binding while intracellular domains can either be linked to intermediate filaments, forming hemidesmosomes, or to actin filaments, forming focal adhesions.[24]

Hemidesmosomes
In hemidesmosomes, integrins attach to extracellular matrix proteins called laminins in the basal lamina, which is the extracellular matrix secreted by epithelial cells.[1] Integrins link extracellular matrix to keratin intermediate filaments, which interacts with intracellular domain of integrins via adapter proteins such as plectins and BP230.[25] Hemidesmosomes are important in maintaining structural stability of epithelial cells by anchoring them together indirectly through the extracellular matrix.
Focal adhesions
In focal adhesions, integrins attach fibronectins, a component in the extracellular matrix, to actin filaments inside cells.[24] Adapter proteins, such as talins, vinculins, α-actinins and filamins, form a complex at the intracellular domain of integrins and bind to actin filaments.[26] This multi-protein complex linking integrins to actin filaments is important for assembly of signalling complexes that act as signals for cell growth and cell motility.[26]
Other organisms
Eukaryotes
Plants cells adhere closely to each other and are connected through plasmodesmata, channels that cross the plant cell walls and connect cytoplasms of adjacent plant cells.[27] Molecules that are either nutrients or signals required for growth are transported, either passively or selectively, between plant cells through plasmodesmata.[27]
Protozoans express multiple adhesion molecules with different specificities that bind to carbohydrates located on surfaces of their host cells.[28] cell–cell adhesion is key for pathogenic protozoans to attach en enter their host cells. An example of a pathogenic protozoan is the malarial parasite (Plasmodium falciparum), which uses one adhesion molecule called the circumsporozoite protein to bind to liver cells,[29] and another adhesion molecule called the merozoite surface protein to bind red blood cells.[30]
Pathogenic fungi use adhesion molecules present on its cell wall to attach, either through protein-protein or protein-carbohydrate interactions, to host cells[31] or fibronectins in the extracellular matrix.[32]
Prokaryotes
Prokaryotes have adhesion molecules on their cell surface termed bacterial adhesins, apart from using its pili (fimbriae) and flagella for cell adhesion.[8] Adhesins can recognise a variety of ligands present on the host cell surfaces and also components in the extracellular matrix. These molecules also control host specificity and regulate tropism (tissue- or cell-specific interactions) through their interaction with their ligands.[33]
Viruses
Viruses also have adhesion molecules required for viral binding to host cells. For example, influenza virus has a hemagglutinin on its surface that is required for recognition of the sugar sialic acid on host cell surface molecules.[34] HIV has an adhesion molecule termed gp120 that binds to its ligand CD4, which is expressed on lymphocytes.[35] Viruses can also target components of cell junctions to enter host cells, which is what happens when the hepatitis C virus targets occludins and claudins in tight junctions to enter liver cells.[9]
Clinical implications
Dysfunction of cell adhesion occurs during cancer metastasis. Loss of cell–cell adhesion in metastatic tumour cells allows them to escape their site of origin and spread through the circulatory system.[5] One example of CAMs deregulated in cancer are cadherins, which are inactivated either by genetic mutations or by other oncogenic signalling molecules, allowing cancer cells to migrate and be more invasive.[6] Other CAMs, like selectins and integrins, can facilitate metastasis by mediating cell–cell interactions between migrating metastatic tumour cells in the circulatory system with endothelial cells of other distant tissues.[36] Due to the link between CAMs and cancer metastasis, these molecules could be potential therapeutic targets for cancer treatment.
There are also other human genetic diseases caused by an inability to express specific adhesion molecules. An example is leukocyte adhesion deficiency-I (LAD-I), where expression of the β2 integrin subunit is reduced or lost.[37] This leads to reduced expression of β2 integrin heterodimers, which are required for leukocytes to firmly attach to the endothelial wall at sites of inflammation in order to fight infections.[38] Leukocytes from LAD-I patients are unable to adhere to endothelial cells and patients exhibit serious episodes of infection that can be life-threatening.
An autoimmune disease called pemphigus is also caused by loss of cell adhesion, as it results from autoantibodies targeting a person's own desmosomal cadherins which leads to epidermal cells detaching from each other and causes skin blistering.[39]
Pathogenic microorganisms, including bacteria, viruses and protozoans, have to first adhere to host cells in order to infect and cause diseases. Anti-adhesion therapy can be used to prevent infection by targeting adhesion molecules either on the pathogen or on the host cell.[40] Apart from altering the production of adhesion molecules, competitive inhibitors that bind to adhesion molecules to prevent binding between cells can also be used, acting as anti-adhesive agents.[41]
See also
References
- Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Morgan, David; Raff, Martin; Roberts, Keith; Walter, Peter (2014). Molecular Biology of the Cell (6th ed.). Garland Science. ISBN 9780815344322.
- Lodish, Harvey; Berk, Arnold; Matsudaira, Paul; Kaiser, Chris A.; Krieger, Monty; Scott, Matthew P.; Zipursky, Lawrence; Darnell, James (2003). Molecular Cell Biology (5th ed.). W.H. Freeman. ISBN 978-0716743668.
- Gumbiner, Barry M. (1996). "Cell Adhesion: The Molecular Basis of Tissue Architecture and Morphogenesis". Cell. 84 (3): 345–357. doi:10.1016/S0092-8674(00)81279-9. PMID 8608588. S2CID 13443584.
- Sumigray, Kaelyn D.; Lechler, Terry (2015). Cell Adhesion in Epidermal Development and Barrier Formation. Current Topics in Developmental Biology. 112. pp. 383–414. doi:10.1016/bs.ctdb.2014.11.027. ISBN 9780124077584. PMC 4737682. PMID 25733147.
- Okegawa, T; Pong, RC; Li, Y; Hsieh, JT (2004). "The role of cell adhesion molecule in cancer progression and its application in cancer therapy". Acta Biochimica Polonica. 51 (2): 445–57. doi:10.18388/abp.2004_3583. PMID 15218541.
- Hirohashi, Setsuo; Kanai, Yae (2003). "Cell adhesion system and human cancer morphogenesis". Cancer Science. 94 (7): 575–581. doi:10.1111/j.1349-7006.2003.tb01485.x. PMID 12841864. S2CID 22154824.
- Szekanecz, Zoltan; Koch, Alisa E (2000). "Cell–cell interactions in synovitis: Endothelial cells and immune cell migration". Arthritis Research. 2 (5): 368–373. doi:10.1186/ar114. PMC 130138. PMID 11094450.
- Pizarro-Cerdá, Javier; Cossart, Pascale (2006). "Bacterial Adhesion and Entry into Host Cells". Cell. 124 (4): 715–727. doi:10.1016/j.cell.2006.02.012. PMID 16497583. S2CID 5769387.
- Mateo, M.; Generous, A.; Sinn, P. L.; Cattaneo, R. (2015). "Connections matter - how viruses use cellcell adhesion components". Journal of Cell Science. 128 (3): 431–439. doi:10.1242/jcs.159400. PMC 4311127. PMID 26046138.
- Meng, W.; Takeichi, M. (2009). "Adherens Junction: Molecular Architecture and Regulation". Cold Spring Harbor Perspectives in Biology. 1 (6): a002899. doi:10.1101/cshperspect.a002899. PMC 2882120. PMID 20457565.
- Nicholl ID, Matsui T, Weiss TM, Stanley CB, Heller WT, Martel A, Farago B, Callaway DJ, Bu Z (Aug 21, 2018). "Alpha-catenin structure and nanoscale dynamics in solution and in complex with F-actin". Biophysical Journal. 115 (4): 642–654. doi:10.1016/j.bpj.2018.07.005. PMC 6104293. PMID 30037495.
- Harris, Tony J. C.; Tepass, Ulrich (2010). "Adherens junctions: from molecules to morphogenesis". Nature Reviews Molecular Cell Biology. 11 (7): 502–514. doi:10.1038/nrm2927. PMID 20571587. S2CID 13638902.
- Johnson, J. L.; Najor, N. A.; Green, K. J. (2014). "Desmosomes: Regulators of Cellular Signaling and Adhesion in Epidermal Health and Disease". Cold Spring Harbor Perspectives in Medicine. 4 (11): a015297. doi:10.1101/cshperspect.a015297. PMC 4208714. PMID 25368015.
- Delva, E.; Tucker, D. K.; Kowalczyk, A. P. (2009). "The Desmosome". Cold Spring Harbor Perspectives in Biology. 1 (2): a002543. doi:10.1101/cshperspect.a002543. PMC 2742091. PMID 20066089.
- Steed, Emily; Balda, Maria S.; Matter, Karl (2010). "Dynamics and functions of tight junctions". Trends in Cell Biology. 20 (3): 142–149. doi:10.1016/j.tcb.2009.12.002. PMID 20061152.
- Niessen, Carien M. (2007). "Tight Junctions/Adherens Junctions: Basic Structure and Function". Journal of Investigative Dermatology. 127 (11): 2525–2532. doi:10.1038/sj.jid.5700865. PMID 17934504.
- Goodenough, D. A.; Paul, D. L. (2009). "Gap Junctions". Cold Spring Harbor Perspectives in Biology. 1 (1): a002576. doi:10.1101/cshperspect.a002576. PMC 2742079. PMID 20066080.
- Meşe, Gülistan; Richard, Gabriele; White, Thomas W. (2007). "Gap Junctions: Basic Structure and Function". Journal of Investigative Dermatology. 127 (11): 2516–2524. doi:10.1038/sj.jid.5700770. PMID 17934503.
- McEver, Rodger P. (2015). "Selectins: initiators of leucocyte adhesion and signalling at the vascular wall". Cardiovascular Research. 107 (3): 331–339. doi:10.1093/cvr/cvv154. PMC 4592324. PMID 25994174.
- Barthel, Steven R; Gavino, Jacyln D; Descheny, Leyla; Dimitroff, Charles J (2007). "Targeting selectins and selectin ligands in inflammation and cancer". Expert Opinion on Therapeutic Targets. 11 (11): 1473–1491. doi:10.1517/14728222.11.11.1473. PMC 2559865. PMID 18028011.
- Wong, Chee Wai; Dye, Danielle E.; Coombe, Deirdre R. (2012). "The Role of Immunoglobulin Superfamily Cell Adhesion Molecules in Cancer Metastasis". International Journal of Cell Biology. 2012: 340296. doi:10.1155/2012/340296. PMC 3261479. PMID 22272201.
- Aricescu, A Radu; Jones, E Yvonne (2007). "Immunoglobulin superfamily cell adhesion molecules: zippers and signals". Current Opinion in Cell Biology. 19 (5): 543–550. doi:10.1016/j.ceb.2007.09.010. PMID 17935964.
- Takada, Yoshikazu; Ye, Xiaojing; Simon, Scott (2007). "The integrins". Genome Biology. 8 (5): 215. doi:10.1186/gb-2007-8-5-215. PMC 1929136. PMID 17543136.
- Lodish, Harvey; Berk, Arnold; Zipursky, S Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James (2000). Molecular cell biology (4th ed.). W.H. Freeman. ISBN 978-0-7167-3136-8.
- Borradori, Luca; Sonnenberg, Arnoud (1999). "Structure and Function of Hemidesmosomes: More Than Simple Adhesion Complexes". Journal of Investigative Dermatology. 112 (4): 411–418. doi:10.1046/j.1523-1747.1999.00546.x. PMID 10201522.
- Critchley, David R (2000). "Focal adhesions – the cytoskeletal connection". Current Opinion in Cell Biology. 12 (1): 133–139. doi:10.1016/S0955-0674(99)00067-8. PMID 10679361.
- Cilia, Michelle Lynn; Jackson, David (2004). "Plasmodesmata form and function". Current Opinion in Cell Biology. 16 (5): 500–506. doi:10.1016/j.ceb.2004.08.002. PMID 15363799.
- Singh, Ram Sarup; Walia, Amandeep Kaur; Kanwar, Jagat Rakesh (2016). "Protozoa lectins and their role in host–pathogen interactions". Biotechnology Advances. 34 (5): 1018–1029. doi:10.1016/j.biotechadv.2016.06.002. PMID 27268207.
- Rathore, Dharmendar; Sacci, John B.; de la Vega, Patricia; McCutchan, Thomas F. (2002). "Binding and Invasion of Liver Cells by Sporozoites". Journal of Biological Chemistry. 277 (9): 7092–7098. doi:10.1074/jbc.M106862200. PMID 11751898.
- Kadekoppala, Madhusudan; Holder, Anthony A. (2010). "Merozoite surface proteins of the malaria parasite: The MSP1 complex and the MSP7 family". International Journal for Parasitology. 40 (10): 1155–1161. doi:10.1016/j.ijpara.2010.04.008. PMID 20451527.
- Tronchin, Guy; Pihet, Marc; Lopes-Bezerra, Leila M.; Bouchara, Jean-Philippe (2008). "Adherence mechanisms in human pathogenic fungi". Medical Mycology. 46 (8): 749–772. doi:10.1080/13693780802206435. PMID 18651303.
- Lima, O. C.; Figueiredo, C. C.; Previato, J. O.; Mendonca-Previato, L.; Morandi, V.; Lopes Bezerra, L. M. (2001). "Involvement of Fungal Cell Wall Components in Adhesion of Sporothrix schenckii to Human Fibronectin". Infection and Immunity. 69 (11): 6874–6880. doi:10.1128/IAI.69.11.6874-6880.2001. PMC 100066. PMID 11598061.
- Klemm, Per; Schembri, Mark A. (2000). "Bacterial adhesins: function and structure". International Journal of Medical Microbiology. 290 (1): 27–35. doi:10.1016/S1438-4221(00)80102-2. PMID 11043979.
- Garman, E. F. (2015). "Antiviral adhesion molecular mechanisms for influenza: W. G. Laver's lifetime obsession". Philosophical Transactions of the Royal Society B: Biological Sciences. 370 (1661): 20140034. doi:10.1098/rstb.2014.0034. PMC 4275904. PMID 25533092.
- Capon, D J; Ward, R H R (1991). "The CD4-gpl20 Interaction and Aids Pathogenesis". Annual Review of Immunology. 9 (1): 649–678. doi:10.1146/annurev.iy.09.040191.003245. PMID 1910691.
- Bendas, Gerd; Borsig, Lubor (2012). "Cancer Cell Adhesion and Metastasis: Selectins, Integrins, and the Inhibitory Potential of Heparins". International Journal of Cell Biology. 2012: 676731. doi:10.1155/2012/676731. PMC 3296185. PMID 22505933.
- Harris, Estelle S.; Weyrich, Andrew S.; Zimmerman, Guy A. (2012). "Lessons from rare maladies: leukocyte adhesion deficiency syndromes". Current Opinion in Hematology. 20 (1): 16–25. doi:10.1097/MOH.0b013e32835a0091. PMC 3564641. PMID 23207660.
- Hanna, Suhair; Etzioni, Amos (2012). "Leukocyte adhesion deficiencies". Annals of the New York Academy of Sciences. 1250 (1): 50–55. Bibcode:2012NYASA1250...50H. doi:10.1111/j.1749-6632.2011.06389.x. PMID 22276660.
- Tamgadge, Sandhya; Bhatt, DaivatM; Pereira, Treville; Tamgadge, Avinash; Bhalerao, Sudhir (2011). "Pemphigus vulgaris". Contemporary Clinical Dentistry. 2 (2): 134–7. doi:10.4103/0976-237X.83074. PMC 3180831. PMID 21957393.
- Krachler, Anne Marie; Orth, Kim (2014). "Targeting the bacteria–host interface". Virulence. 4 (4): 284–294. doi:10.4161/viru.24606. PMC 3710331. PMID 23799663.
- Ofek, Itzhak; Hasty, David L; Sharon, Nathan (2003). "Anti-adhesion therapy of bacterial diseases: prospects and problems". FEMS Immunology & Medical Microbiology. 38 (3): 181–191. CiteSeerX 10.1.1.320.1480. doi:10.1016/S0928-8244(03)00228-1. PMID 14522453.
External links
| Wikimedia Commons has media related to Cell adhesion. |
- The Cell by G. Cooper (online textbook)
- Molecular Cell Biology by Lodish et al. (online textbook)
- Molecular Biology of the Cell by Alberts et al. (online textbook)
- Cell Adhesion and Extracellular Matrix - The Virtual Library of Biochemistry, Molecular Biology and Cell Biology