Defensin
Defensins are small cysteine-rich cationic proteins across cellular life, including vertebrate[1] and invertebrate[2] animals, plants,[3][4] and fungi.[5] They are host defense peptides, with members displaying either direct antimicrobial activity, immune signalling activities, or both. They are variously active against bacteria, fungi and many enveloped and nonenveloped viruses. They are typically 18-45 amino acids in length, with three or four highly conserved disulphide bonds.
| Defensin | |
|---|---|
 | |
| Identifiers | |
| Symbol | Defensin |
| Pfam clan | CL0075 |
| OPM superfamily | 54 |
| OPM protein | 6cs9 |
In animals, they are produced by cells of the innate immune system and epithelial cells, whereas in plants and fungi they are produced by a wide variety of tissues. An organism usually produces many different defensins, some of which are stored inside the cells (e.g. in neutrophil granulocytes to kill phagocytosed bacteria), and others are secreted into the extracellular medium. For those that directly kill microbes, their mechanism of action varies from disruption of the microbial cell membrane to metabolic disruption.
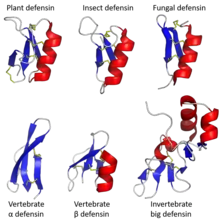
Varieties
The name 'defensin' was coined in the mid 1980s, though the proteins have been variously called 'Cationic Antimicrobial Proteins', 'Neutrophil peptides', 'Gamma thionins' amongst others.[6]
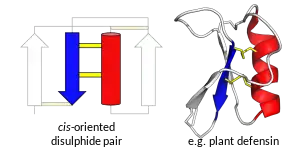
Proteins called 'defensins' are not all evolutionarily related to one another.[7] Instead fall into two broad superfamilies, each of which contains multiple families.[7][8] One superfamily, the trans-defensins, contains the defensins found in humans and other vertebrates[9][10] as well as some invertebrates.[11][12] The other superfamily, cis-defensins, contains the defensins found in invertebrates, plants and fungi.[13][14][15] The superfamilies and families are determined overall tertiary structure and each family usually has a conserved pattern of disulphide bonds.[9][16] All defensins form small and compact folded structures, typically with a high positive charge that are highly stable due to the multiple disulphide bonds. In all families, the underlying genes responsible for defensin production are highly polymorphic.
Trans-defensins
Vertebrate defensins are primarily α-defensins and the β-defensins. Some primates additionally have the much smaller θ-defensins. In general, both α- and β-defensins are encoded by two-exon genes, where the first exon encodes for a hydrophobic leader sequence (that is removed after translation) and the cysteine-rich sequence (the mature peptide). The disulfide linkages formed by the cysteines have been suggested to be essential for activities related to innate immunity in mammals, but are not necessarily required for antimicrobial activity.[17][18] Theta defensins form a single beta hairpin structure and therefore also represent a distinct group. Only alpha and beta defensins are expressed in humans.[19]
Although the most well studied defensins are from vertebrates, a family of trans-defensins called 'big defensins' are found in molluscs, arthropods and lancelets.[7][8]
Cis-defensins
Arthropod defensins are the best-characterised defensins from invertebrates (especially those from insects).[23] Other invertebrates known to produce defensins from this protein superfamily include molluscs, annelids and cnidaria.[24]
Plants defensins were discovered in 1990 and have subsequently been found in most plant tissues with antimicrobial activities, with both antifungal and antibacterial examples.[25] They have been identified in all major groups of vascular plants, but not in ferns, mosses or algae.[25]
Fungal defensins were first identified in 2005.[26] Studied examples mainly have anti-bacterial activities and have been found in both main divisions of fungi (Ascomycota and Basidiomycota), as well as in the more basal groups of Zygomycota and Glomeromycota.[27]
Bacterial defensins have also been identified, but are by far the least studied. They include variants with only four cysteines, whereas defensins from eukaryote defensins almost all have six or eight.[28]
Related defensin-like proteins
In addition to the defensins involved in host defence, there are a number of related Defensin-Like Peptides (DLPs) that have evolved to have other activities.
Toxins
There appear to have been multiple evolutionary recruitments of defensins to be toxin proteins used in the venoms of animals.[29] and act via a completely different mechanism to their antimicrobial relatives, from binding directly to ion channels to disrupting nerve signals. Examples include the crotamine toxin in snake venom,[30] many scorpion toxins,[31] some sea anemone toxins,[10] and one of the toxins in platypus venom.[29] Indeed, an insect defensin has been experimentally converted into a toxin by deletion of a small loop that otherwise sterically hindered interactions with the ion channels.[32]
Signalling
In vertebrates, some α- and β-defensins are involved in signalling between the innate immune and adaptive immune systems.[33][34] In plants, a specialised family of DLPs is involved in signalling to detect if self-pollination has occurred and induce self-incompatibility to prevent inbreeding.[35]
Enzyme inhibitors
Some antimicrobial defensins also have enzyme inhibitory activity, and some DLPs function primarily as enzyme inhibitors, acting as antifeedants (discouraging animals from eating them).[36][37][38]
Function
In immature marsupials, because their immune system is underdeveloped at the time of birth, defensins play a major role in defense against pathogens. They are produced in the milk of the mother as well as by the young marsupial in question.
In human breast milk, defensins play a central role in neonate immunity.[39]
The human genome contains theta-defensin genes, but they have a premature stop codon, hampering their expression. An artificial human theta-defensin,[40] retrocyclin, was created by 'fixing' the pseudogene, and it was shown to be effective against HIV[41] and other viruses, including herpes simplex virus and influenza A. They act primarily by preventing these viruses from entering their target cells.
Also interesting is the effect of alpha-defensins on the exotoxin produced by anthrax (Bacillus anthracis). Chun Kim et al. showed how anthrax, which produces a metalloprotease lethal factor (LF) protein to target MAPKK, is vulnerable to human neutrophil protein-1 (HNP-1). This group showed HNP-1 to behave as a reversible noncompetitive inhibitor of LF.[42]
They have generally been considered to contribute to mucosal health; however, it is possible that these peptides can be considered biological factors that can be upregulated by bioactive compounds present in human breast milk. In this sense, the intestinal production of antimicrobial peptides as hBD2 and hBD4 by trefoil from milk might play an important role on neonate colonization, thereby enhancing the immune response of newborns against pathogens with which they may come in contact.[39][43]
Pathology
The alpha defensin peptides are increased in chronic inflammatory conditions.
Alpha defensin are increased in several cancers, including colorectal cancer.[44]
An imbalance of defensins in the skin may contribute to acne.[45]
A reduction of ileal defensins may predispose to Crohn's disease.[46][47]
In one small study, a significant increase in alpha defensin levels was detected in T cell lysates of schizophrenia patients; in discordant twin pairs, unaffected twins also had an increase, although not as high as that of their ill siblings. The authors suggested that alpha-defensin levels might prove a useful marker for schizophrenia risk.[48]
Defensins are found in the human skin during inflammatory conditions like psoriasis[49] and also during wound healing.
Applications
Defensins
At present, the widespread spread of antibiotic resistance requires the search and development of new antimicrobial drugs. From this point of view, defensins (as well as antimicrobial peptides in general) are of great interest. It was shown that defensins have pronounced antibacterial activity against a wide range of pathogens.[50] In addition, defensins can enhance the effectiveness of conventional antibiotics.[50]
Defensin-mimetics
Defensin mimetics, also called host defense peptide (HDP) mimetics, are completely synthetic, non-peptide, small molecule structures that mimic defensins in structure and activity.[51] Similar molecules, such as brilacidin, are being developed as antibiotics,[52] anti-inflammatories for oral mucositis,[53][54] and antifungals, especially for candidiasis.[55][56][57]
See also
- Host defense peptides, to which defensins belong
References
- Hazlett L, Wu M (January 2011). "Defensins in innate immunity". Cell and Tissue Research. 343 (1): 175–88. doi:10.1007/s00441-010-1022-4. PMID 20730446. S2CID 2234617.
- Tassanakajon A, Somboonwiwat K, Amparyup P (February 2015). "Sequence diversity and evolution of antimicrobial peptides in invertebrates". Developmental and Comparative Immunology. Specific immunity in invertebrates. 48 (2): 324–41. doi:10.1016/j.dci.2014.05.020. PMID 24950415.
- Thomma BP, Cammue BP, Thevissen K (December 2002). "Plant defensins". Planta. 216 (2): 193–202. doi:10.1007/s00425-002-0902-6. PMID 12447532. S2CID 19356421.
- Sathoff AE, Samac DA (May 2019). "Antibacterial Activity of Plant Defensins". Molecular Plant-Microbe Interactions. 32 (5): 507–514. doi:10.1094/mpmi-08-18-0229-cr. PMID 30501455.
- Wu J, Gao B, Zhu S (August 2014). "The fungal defensin family enlarged". Pharmaceuticals. 7 (8): 866–80. doi:10.3390/ph7080866. PMC 4165938. PMID 25230677.
- Lehrer RI (September 2004). "Primate defensins". Nature Reviews. Microbiology. 2 (9): 727–38. doi:10.1038/nrmicro976. PMID 15372083. S2CID 8774156.
- Shafee TM, Lay FT, Hulett MD, Anderson MA (September 2016). "The Defensins Consist of Two Independent, Convergent Protein Superfamilies". Molecular Biology and Evolution. 33 (9): 2345–56. doi:10.1093/molbev/msw106. PMID 27297472.
- Shafee TM, Lay FT, Phan TK, Anderson MA, Hulett MD (February 2017). "Convergent evolution of defensin sequence, structure and function". Cellular and Molecular Life Sciences. 74 (4): 663–682. doi:10.1007/s00018-016-2344-5. PMID 27557668. S2CID 24741736.
- Hollox EJ, Abujaber R (2017). "Evolution and Diversity of Defensins in Vertebrates". In Pontarotti P (ed.). Evolutionary Biology: Self/Nonself Evolution, Species and Complex Traits Evolution, Methods and Concepts. Springer International Publishing. pp. 27–50. doi:10.1007/978-3-319-61569-1_2. ISBN 978-3-319-61569-1.
- Mitchell ML, Shafee T, Papenfuss AT, Norton RS (July 2019). "Evolution of cnidarian trans-defensins: Sequence, structure and exploration of chemical space". Proteins. 87 (7): 551–560. doi:10.1002/prot.25679. PMID 30811678. S2CID 73469576.
- Zhu S, Gao B (2013). "Evolutionary origin of β-defensins". Developmental and Comparative Immunology. 39 (1–2): 79–84. doi:10.1016/j.dci.2012.02.011. PMID 22369779.
- Montero-Alejo V, Corzo G, Porro-Suardíaz J, Pardo-Ruiz Z, Perera E, Rodríguez-Viera L, et al. (February 2017). "Panusin represents a new family of β-defensin-like peptides in invertebrates". Developmental and Comparative Immunology. 67: 310–321. doi:10.1016/j.dci.2016.09.002. PMID 27616720. S2CID 19734223.
- Dias RD, Franco OL (October 2015). "Cysteine-stabilized αβ defensins: From a common fold to antibacterial activity". Peptides. Festschrift to highlight the career of Abba J. Kastin as a founding editor, researcher, and educator in the peptide field. 72: 64–72. doi:10.1016/j.peptides.2015.04.017. PMID 25929172. S2CID 17846143.
- Shafee T, Anderson MA (March 2019). "A quantitative map of protein sequence space for the cis-defensin superfamily". Bioinformatics. 35 (5): 743–752. doi:10.1093/bioinformatics/bty697. PMID 30102339. S2CID 51968286.
- Zhu S (February 2008). "Discovery of six families of fungal defensin-like peptides provides insights into origin and evolution of the CSalphabeta defensins". Molecular Immunology. 45 (3): 828–38. doi:10.1016/j.molimm.2007.06.354. PMID 17675235.
- Wang YP, Lai R (February 2010). "[Insect antimicrobial peptides: structures, properties and gene regulation]". Dong Wu Xue Yan Jiu = Zoological Research. 31 (1): 27–34. doi:10.3724/sp.j.1141.2010.01027. PMID 20446450.
- Varkey J, Singh S, Nagaraj R (November 2006). "Antibacterial activity of linear peptides spanning the carboxy-terminal beta-sheet domain of arthropod defensins". Peptides. 27 (11): 2614–23. doi:10.1016/j.peptides.2006.06.010. PMID 16914230. S2CID 21104756.
- Varkey J, Nagaraj R (November 2005). "Antibacterial activity of human neutrophil defensin HNP-1 analogs without cysteines". Antimicrobial Agents and Chemotherapy. 49 (11): 4561–6. doi:10.1128/AAC.49.11.4561-4566.2005. PMC 1280114. PMID 16251296.
- Dhople V, Krukemeyer A, Ramamoorthy A (September 2006). "The human beta-defensin-3, an antibacterial peptide with multiple biological functions". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1758 (9): 1499–512. doi:10.1016/j.bbamem.2006.07.007. PMID 16978580.
- Tran D, Tran P, Roberts K, Osapay G, Schaal J, Ouellette A, Selsted ME (March 2008). "Microbicidal properties and cytocidal selectivity of rhesus macaque theta defensins". Antimicrobial Agents and Chemotherapy. 52 (3): 944–53. doi:10.1128/AAC.01090-07. PMC 2258523. PMID 18160518.
- Garcia AE, Selsted M (March 2008). "Olive baboon θ-defensins". The FASEB Journal. 22 (1 Suppl): 673.11. doi:10.1096/fasebj.22.1_supplement.673.11 (inactive 2021-01-10).CS1 maint: DOI inactive as of January 2021 (link)
- Garcia AE, Osapay G, Tran PA, Yuan J, Selsted ME (December 2008). "Isolation, synthesis, and antimicrobial activities of naturally occurring theta-defensin isoforms from baboon leukocytes". Infection and Immunity. 76 (12): 5883–91. doi:10.1128/IAI.01100-08. PMC 2583559. PMID 18852242.
- Koehbach J (2017). "Structure-Activity Relationships of Insect Defensins". Frontiers in Chemistry. 5: 45. Bibcode:2017FrCh....5...45K. doi:10.3389/fchem.2017.00045. PMC 5506212. PMID 28748179.
- Greco S, Gerdol M, Edomi P, Pallavicini A (January 2020). "Molecular Diversity of Mytilin-Like Defense Peptides in Mytilidae (Mollusca, Bivalvia)". Antibiotics. 9 (1): 37. doi:10.3390/antibiotics9010037. PMC 7168163. PMID 31963793.
- Parisi K, Shafee TM, Quimbar P, van der Weerden NL, Bleackley MR, Anderson MA (April 2019). "The evolution, function and mechanisms of action for plant defensins". Seminars in Cell & Developmental Biology. 88: 107–118. doi:10.1016/j.semcdb.2018.02.004. PMID 29432955.
- Mygind PH, Fischer RL, Schnorr KM, Hansen MT, Sönksen CP, Ludvigsen S, et al. (October 2005). "Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus". Nature. 437 (7061): 975–80. Bibcode:2005Natur.437..975M. doi:10.1038/nature04051. PMID 16222292. S2CID 4423851.
- Wu J, Gao B, Zhu S (August 2014). "The fungal defensin family enlarged". Pharmaceuticals. 7 (8): 866–80. doi:10.3390/ph7080866. PMC 4165938. PMID 25230677.
- Dash TS, Shafee T, Harvey PJ, Zhang C, Peigneur S, Deuis JR, et al. (February 2019). "A Centipede Toxin Family Defines an Ancient Class of CSαβ Defensins". Structure. 27 (2): 315–326.e7. doi:10.1016/j.str.2018.10.022. PMID 30554841.
- Whittington CM, Papenfuss AT, Bansal P, Torres AM, Wong ES, Deakin JE, et al. (June 2008). "Defensins and the convergent evolution of platypus and reptile venom genes". Genome Research. 18 (6): 986–94. doi:10.1101/gr.7149808. PMC 2413166. PMID 18463304.
- Batista da Cunha D, Pupo Silvestrini AV, Gomes da Silva AC, Maria de Paula Estevam D, Pollettini FL, de Oliveira Navarro J, et al. (May 2018). "Mechanistic insights into functional characteristics of native crotamine". Toxicon. 146: 1–12. doi:10.1016/j.toxicon.2018.03.007. hdl:11449/170828. PMID 29574214. S2CID 205440053.
- Possani LD, Becerril B, Delepierre M, Tytgat J (September 1999). "Scorpion toxins specific for Na+-channels". European Journal of Biochemistry. 264 (2): 287–300. doi:10.1046/j.1432-1327.1999.00625.x. PMID 10491073.
- Zhu S, Peigneur S, Gao B, Umetsu Y, Ohki S, Tytgat J (March 2014). "Experimental conversion of a defensin into a neurotoxin: implications for origin of toxic function". Molecular Biology and Evolution. 31 (3): 546–59. doi:10.1093/molbev/msu038. PMID 24425781.
- Petrov V, Funderburg N, Weinberg A, Sieg S (December 2013). "Human β defensin-3 induces chemokines from monocytes and macrophages: diminished activity in cells from HIV-infected persons". Immunology. 140 (4): 413–20. doi:10.1111/imm.12148. PMC 3839645. PMID 23829433.
- Semple F, Dorin JR (2012). "β-Defensins: multifunctional modulators of infection, inflammation and more?". Journal of Innate Immunity. 4 (4): 337–48. doi:10.1159/000336619. PMC 6784047. PMID 22441423.
- Fobis-Loisy I, Ivanov R, Gaude T (2012). "The S-LOCUS CYSTEINE-RICH PROTEIN (SCR): A Small Peptide with A High Impact on the Evolution of Flowering Plants". Plant Signaling Peptides. Signaling and Communication in Plants. 16. Springer Berlin Heidelberg. pp. 77–92. doi:10.1007/978-3-642-27603-3_5. ISBN 978-3-642-27602-6.
- Williams LK, Brayer GD (2015-11-25). "Porcine pancreatic alpha-amylase in complex with helianthamide, a novel proteinaceous inhibitor". doi:10.2210/pdb4x0n/pdb.
- Zhao Q, Chae YK, Markley JL (2003-01-07). "Minimized NMR structure of ATT, an Arabidopsis trypsin/chymotrypsin inhibitor". doi:10.2210/pdb1jxc/pdb.
- Pelegrini PB, Lay FT, Murad AM, Anderson MA, Franco OL (November 2008). "Novel insights on the mechanism of action of alpha-amylase inhibitors from the plant defensin family". Proteins. 73 (3): 719–29. doi:10.1002/prot.22086. PMID 18498107. S2CID 28378146.
- Barrera GJ, Sanchez G, Gonzalez JE (November 2012). "Trefoil factor 3 isolated from human breast milk downregulates cytokines (IL8 and IL6) and promotes human beta defensin (hBD2 and hBD4) expression in intestinal epithelial cells HT-29". Bosnian Journal of Basic Medical Sciences. 12 (4): 256–64. doi:10.17305/bjbms.2012.2448. PMC 4362502. PMID 23198942.
- retrocyclin at the US National Library of Medicine Medical Subject Headings (MeSH)
- Münk C, Wei G, Yang OO, Waring AJ, Wang W, Hong T, et al. (October 2003). "The theta-defensin, retrocyclin, inhibits HIV-1 entry". AIDS Research and Human Retroviruses. 19 (10): 875–81. doi:10.1089/088922203322493049. PMID 14585219.
- Kim C, Gajendran N, Mittrücker HW, Weiwad M, Song YH, Hurwitz R, et al. (March 2005). "Human alpha-defensins neutralize anthrax lethal toxin and protect against its fatal consequences". Proceedings of the National Academy of Sciences of the United States of America. 102 (13): 4830–5. Bibcode:2005PNAS..102.4830K. doi:10.1073/pnas.0500508102. PMC 555714. PMID 15772169.
- Barrera GJ, Tortolero GS (2016). "Trefoil factor 3 (TFF3) from human breast milk activates PAR-2 receptors, of the intestinal epithelial cells HT-29, regulating cytokines and defensins". Bratislavske Lekarske Listy. 117 (6): 332–9. doi:10.4149/bll_2016_066. PMID 27546365.
- Albrethsen J, Bøgebo R, Gammeltoft S, Olsen J, Winther B, Raskov H (January 2005). "Upregulated expression of human neutrophil peptides 1, 2 and 3 (HNP 1-3) in colon cancer serum and tumours: a biomarker study". BMC Cancer. 5: 8. doi:10.1186/1471-2407-5-8. PMC 548152. PMID 15656915.
- Philpott MP (November 2003). "Defensins and acne". Molecular Immunology. 40 (7): 457–62. doi:10.1016/S0161-5890(03)00154-8. PMID 14568392.
- "Researchers discover a possible cause of chronic inflammations of Crohn Disease". Genomics & Genetics Weekly: 72. August 11, 2006.
- Wehkamp J, Salzman NH, Porter E, Nuding S, Weichenthal M, Petras RE, et al. (December 2005). "Reduced Paneth cell alpha-defensins in ileal Crohn's disease". Proceedings of the National Academy of Sciences of the United States of America. 102 (50): 18129–34. Bibcode:2005PNAS..10218129W. doi:10.1073/pnas.0505256102. PMC 1306791. PMID 16330776.
- Craddock RM, Huang JT, Jackson E, Harris N, Torrey EF, Herberth M, Bahn S (July 2008). "Increased alpha-defensins as a blood marker for schizophrenia susceptibility". Molecular & Cellular Proteomics. 7 (7): 1204–13. doi:10.1074/mcp.M700459-MCP200. PMID 18349140. S2CID 35381828.
- Harder J, Bartels J, Christophers E, Schroder JM (February 2001). "Isolation and characterization of human beta -defensin-3, a novel human inducible peptide antibiotic". The Journal of Biological Chemistry. 276 (8): 5707–13. doi:10.1074/jbc.M008557200. PMID 11085990. S2CID 9516726.
- Bolatchiev A (2020-11-25). "Antibacterial activity of human defensins against Staphylococcus aureus and Escherichia coli". PeerJ. 8: e10455. doi:10.7717/peerj.10455. PMC 7698690. PMID 33304659.
- "Press release: PolyMedix". 2008-05-09. Business Wire
- "PMX-30063 The First And Only Defensin Mimetic Systemic Antibiotic Drug In Human Clinical Trials". 2008.
- Clinical trial number NCT02324335 for "Phase 2 Study to Evaluate the Safety & Efficacy of Brilacidin Oral Rinse in Patients With Head and Neck Cancer (Brilacidin)" at ClinicalTrials.gov
- "Brilacidin-OM page". Cellceutix. Archived from the original on 2015-02-07. Retrieved 2015-03-02.
- "Candidiasis". Cellceutix. Archived from the original on 2015-02-07. Retrieved 2015-03-02.
- Diamond G, Scott R. "A Novel Therapeutic For Invasive Candiasis". Grantome. Fox Chase Chemical Diversity Center.
- Ryan LK, Freeman KB, Masso-Silva JA, Falkovsky K, Aloyouny A, Markowitz K, et al. (July 2014). "Activity of potent and selective host defense peptide mimetics in mouse models of oral candidiasis". Antimicrobial Agents and Chemotherapy. 58 (7): 3820–7. doi:10.1128/AAC.02649-13. PMC 4068575. PMID 24752272.
External links
| Wikimedia Commons has media related to Defensin. |
- Defensins Database, Singapore
- Innate ( Nonspecific ) Immunity at Western Kentucky University
- UMich Orientation of Proteins in Membranes families/superfamily-56 - Vertebrate defensins and related sea anemone sodium channel toxins
- UMich Orientation of Proteins in Membranes families/superfamily-61 - Defensins from insects and plants and scorpion toxins
- Defensins at the US National Library of Medicine Medical Subject Headings (MeSH)