Glyoxylate reductase
Glyoxylate reductase (EC 1.1.1.26), first isolated from spinach leaves,[2] is an enzyme that catalyzes the reduction of glyoxylate to glycolate, using the cofactor NADH or NADPH.
| Glyoxylate Reductase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
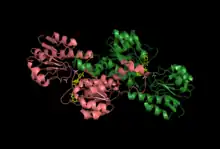 The crystal structure of glyoxylate reductase complexed with NADPH. Each color represents a monomer of the enzyme. This enzyme was isolated from a hyperthermophilic archaeon, Pyrococcus horikoshii OT3.[1]
Image created by PyMOL. | |||||||||
| Identifiers | |||||||||
| EC number | 1.1.1.26 | ||||||||
| CAS number | 9028-32-4 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
The systematic name of this enzyme class is glycolate:NAD+ oxidoreductase. Other names in common use include NADH-glyoxylate reductase, glyoxylic acid reductase, and NADH-dependent glyoxylate reductase.
Structure
The crystal structure of the glyoxylate reductase enzyme from the hyperthermophilic archeon Pyrococcus horiskoshii OT3 has been reported.[1] The enzyme exists in the dimeric form. Each monomer has two domains: a substrate-binding domain where glyoxylate binds, and a nucleotide-binding domain where the NAD(P)H cofactor binds.
Mechanism
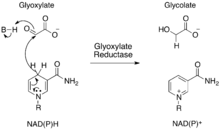
The enzyme catalyzes the transfer of a hydride from NAD(P)H to glyoxylate, causing a reduction of the substrate to glycolate and an oxidation of the cofactor to NAD(P)+. Figure 2 shows the mechanism for this reaction.
It is thought that the two residues Glu270 and His288 are important for the enzyme's catalytic function, while the residue Arg241 is thought to be important for substrate specificity.[1]
Function
The glyoxylate reductase enzyme localizes to the cell cytoplasm in plants. It can use both NADPH and NADH as a cofactor, but prefers NADPH. The enzyme substrate, glyoxylate, is a metabolite in plant photorespiration, and is produced in the peroxisome. Glyoxylate is important in the plant cell as it can deactivate RUBISCO and inhibit its activation. Hence, glyoxylate levels are important in regulating photosynthesis.[3]
The enzyme is thought of as a glyoxylate-glycolate shuttle that helps in the disposal of excess reducing equivalents from photosynthesis. This is supported by the following findings: (1) glycolate biosynthesis in the chloroplasts is highest at low CO2 concentrations, (2) the enzyme is quite specific for the NADPH cofactor which is a final product of electron transfer in the chloroplasts during photosynthesis, and (3) when isolated chloroplasts are exposed to light, they absorb glyoxylate and reduce it, but they do not absorb glycolate.[4]
Due to the link between glyoxylate levels and photosynthesis, an increase in glyoxylate levels indicates that the plant is under stress. As glyoxylate levels continue to increase, they can harm the plant by (1) reacting with DNA, (2) oxidizing membrane lipids, (3) modifying proteins, and (4) increasing the transcription of stress-related genes in the plant. This highlights the importance of glyoxylate reductase, as it helps keep plant cells healthy and detoxifies the cell by reducing glyoxylate levels. In the absence of the enzyme, the side-effects of increased glyoxylate activity can cause cellular and developmental problems in the plant.[5]
Glyoxylate reductase can be used as a tool for studying photorespiratory carbon metabolism in plant leaves. Such studies can be carried out using acetohydroxamate and aminooxyacetate, which have been found to inhibit glyoxylate reductase activity. These inhibitors are not fully specific, but provide fully reversible inhibition of the enzyme and so provide a flexible tool for metabolic studies in plants.[6]
Disease relevance
A human protein, GRHPR, has been identified that exhibits both glyoxylate and hydroxypyruvate reductase activities. The DNA sequence of this protein is up to 30% similar to the sequence of hydroxypyruvate and glyoxylate reductases found in a range of plant and microbial species.[7]
GRHPR is an important protein in the human body, as it converts the metabolic byproduct glyoxylate into the less reactive glycolate.[8] The reduced function of the enzyme causes a build-up of glyoxylate in the liver, and in turn causes an increase in oxalate levels in urine.[9]
The reduced enzyme function can be caused by a rare inherited autosomal recessive[10] disorder known as primary hyperoxaluria type II (PH2). This condition can cause nephrolithiasis (kidney stone), nephrocalcinosis and renal failure.[11][12]
Industrial relevance
Glyoxylate reductase uses NAD(P)H to reduce an oxoacid (glyoxylate) to its corresponding α-hydroxy acid (glycolate). This class of reactions provides an opportunity for the synthesis of chiral hydroxy acids. Such products are of interest in the synthesis of pharmaceuticals, such as anti-obesity compounds and semisynthetic penicillins.[13]
Evolution
Glyoxylate is an important component of the glyoxylate cycle, a variant of the citric acid cycle, whereby acetyl-CoA is converted to succinate and then other carbohydrates in plants, bacteria, protists, and fungi. Studies have been conducted to trace the genes for the glyoxylate cycle enzymes to animals. The studies have shown that these genes are in fact present in animals, but the redistribution of the genes suggest that either that (1) these genes encode other enzymes that take part in the glyoxylate cycle, but are not orthologous to the known enzymes in the cycle, or (2) animals have developed a new function for these enzymes that have yet to be characterized.[14]
References
- Yoshikawa S, Arai R, Kinoshita Y, Uchikubo-Kamo T, Wakamatsu T, Akasaka R, Masui R, Terada T, Kuramitsu S, Shirouzu M, Yokoyama S (March 2007). "Structure of archaeal glyoxylate reductase from Pyrococcus horikoshii OT3 complexed with nicotinamide adenine dinucleotide phosphate". Acta Crystallogr. D. 63 (Pt 3): 357–65. doi:10.1107/S0907444906055442. PMID 17327673.
- ZELITCH I (April 1953). "Oxidation and reduction of glycolic and glyoxylic acids in plants. II. Glyoxylic acid reductase". J. Biol. Chem. 201 (2): 719–26. PMID 13061410.
- Givan CV, Kleczkowski LA (October 1992). "The enzymic reduction of glyoxylate and hydroxypyruvate in leaves of higher plants". Plant Physiol. 100 (2): 552–6. doi:10.1104/pp.100.2.552. PMC 1075593. PMID 16653027.
- Tolbert NE, Yamazaki RK, Oeser A (October 1970). "Localization and properties of hydroxypyruvate and glyoxylate reductases in spinach leaf particles". J. Biol. Chem. 245 (19): 5129–36. PMID 4394164.
- Allan WL, Clark SM, Hoover GJ, Shelp BJ (October 2009). "Role of plant glyoxylate reductases during stress: a hypothesis". Biochem. J. 423 (1): 15–22. doi:10.1042/BJ20090826. PMC 2762691. PMID 19740079.
- Kleczkowski LA, Randall DD, Blevins DG (July 1987). "Inhibition of Spinach Leaf NADPH(NADH)-Glyoxylate Reductase by Acetohydroxamate, Aminooxyacetate, and Glycidate". Plant Physiol. 84 (3): 619–23. doi:10.1104/pp.84.3.619. PMC 1056639. PMID 16665491.
- Rumsby G, Cregeen DP (September 1999). "Identification and expression of a cDNA for human hydroxypyruvate/glyoxylate reductase". Biochim. Biophys. Acta. 1446 (3): 383–8. doi:10.1016/S0167-4781(99)00105-0. PMID 10524214.
- Mdluli K, Booth MP, Brady RL, Rumsby G (December 2005). "A preliminary account of the properties of recombinant human Glyoxylate reductase (GRHPR), LDHA and LDHB with glyoxylate, and their potential roles in its metabolism". Biochim. Biophys. Acta. 1753 (2): 209–16. doi:10.1016/j.bbapap.2005.08.004. PMID 16198644.
- Booth MP, Conners R, Rumsby G, Brady RL (June 2006). "Structural basis of substrate specificity in human glyoxylate reductase/hydroxypyruvate reductase". J. Mol. Biol. 360 (1): 178–89. doi:10.1016/j.jmb.2006.05.018. PMID 16756993.
- Cregeen DP, Williams EL, Hulton S, Rumsby G (December 2003). "Molecular analysis of the glyoxylate reductase (GRHPR) gene and description of mutations underlying primary hyperoxaluria type 2". Hum. Mutat. 22 (6): 497. doi:10.1002/humu.9200. PMID 14635115. S2CID 39645821.
- Cramer SD, Ferree PM, Lin K, Milliner DS, Holmes RP (October 1999). "The gene encoding hydroxypyruvate reductase (GRHPR) is mutated in patients with primary hyperoxaluria type II". Hum. Mol. Genet. 8 (11): 2063–9. doi:10.1093/hmg/8.11.2063. PMID 10484776.
- Lam CW, Yuen YP, Lai CK, Tong SF, Lau LK, Tong KL, Chan YW (December 2001). "Novel mutation in the GRHPR gene in a Chinese patient with primary hyperoxaluria type 2 requiring renal transplantation from a living related donor". Am. J. Kidney Dis. 38 (6): 1307–10. doi:10.1053/ajkd.2001.29229. PMID 11728965.
- Hummel W, Kula MR (September 1989). "Dehydrogenases for the synthesis of chiral compounds". Eur. J. Biochem. 184 (1): 1–13. doi:10.1111/j.1432-1033.1989.tb14983.x. PMID 2673781.
- Kondrashov FA, Koonin EV, Morgunov IG, Finogenova TV, Kondrashova MN (2006). "Evolution of glyoxylate cycle enzymes in Metazoa: evidence of multiple horizontal transfer events and pseudogene formation". Biol. Direct. 1: 31. doi:10.1186/1745-6150-1-31. PMC 1630690. PMID 17059607.