HLA-A36
HLA-A36 (A36) is a human leukocyte antigen serotype within HLA-A serotype group. The serotype is determined by the antibody recognition of α36 subset of HLA-A α-chains. For A36, the alpha "A" chain are encoded by the HLA-A*36 allele group and the β-chain are encoded by B2M locus.[1] This group currently is dominated by A*3601. A36 and A*36 are almost synonymous in meaning.
| HLA-A36 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (MHC Class I, A cell surface antigen) | ||||||||||||||||
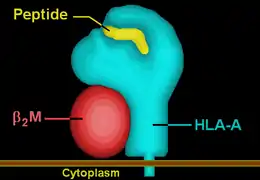 HLA-A36 | ||||||||||||||||
| About | ||||||||||||||||
| Protein | transmembrane receptor/ligand | |||||||||||||||
| Structure | αβ heterodimer | |||||||||||||||
| Subunits | HLA-A*3601, β2-microglobulin | |||||||||||||||
| Subtypes | ||||||||||||||||
| ||||||||||||||||
| Alleles link-out to IMGT/HLA database at EBI | ||||||||||||||||
A36 is more common in Africa (North and East) and Central Asia. A36 is rare HLA-A allele group.
Serotype
| A*36 | A36 | A1 | Sample |
| allele | % | % | size (N) |
| *3601 | 68 | 16 | 807 |
A36 has a high false serotyping rate to A1.
Distribution
| freq | ||||
| ref. | Population | (%) | ||
| Kenya | 6.6 | |||
| [3] | N. African (non-caucasian) | 4.5 | ||
| [4] | Mossi(Burkino Faso) | 3.8 | ||
| [5] | Beti(Cameroon) | 3.7 | ||
| [3] | Mongolian | 3.1 | ||
| Lusaka (Zambia) | 3.1 | |||
| [3] | Harare Shona (Zimbabwe) | 2.9 | ||
| [3] | Uygars (China) | 2.6 | ||
| [5] | Bamileke(Cameroon) | 2.6 | ||
| [6] | Amman (Jordon) | 2.4 | ||
| [4] | Fulani | 2.0 | ||
| [3] | Sweden | 1.6 | ||
| Tswana (S. Africa) | 1.2 | |||
| Kampala (Uganda) | 1.2 | |||
| [3] | Spain | 0.9 | ||
| Bubi(Eq. Guinea) | 0.5 | |||
| [3] | !kung (San) | 0.0 | ||
| Berber(Morocco) | 0.0 |
A36 is largely limited to Africa. Outside Africa more than half of the populations have no A36 and the majority that do, have only trace levels. The exception is in Central/East Central Asia. This appears not to be coincidental. HLA DR3-DQ2 linkage and frequency indicates a strong possibility of a recent migration from either North or West Africa in which the DR3-DQ2 (notably DRB1*0302) and A*33-B*58 haplotypes were carried into and redistributed from Central Asia into the surrounding populations.
References
- Arce-Gomez B, Jones EA, Barnstable CJ, Solomon E, Bodmer WF (February 1978). "The genetic control of HLA-A and B antigens in somatic cell hybrids: requirement for beta2 microglobulin". Tissue Antigens. 11 (2): 96–112. doi:10.1111/j.1399-0039.1978.tb01233.x. PMID 77067.
- Allele Query Form IMGT/HLA - European Bioinformatics Institute
- Sasazuki, Takehiko; Tsuji, Kimiyoshi; Aizawa, Miki (1992). HLA 1991: proceedings of the eleventh International Histocompatibility Workshop and Conference, held in Yokohama, Japan, 6–13 November 1991. Oxford [Oxfordshire]: Oxford University Press. ISBN 0-19-262390-7.
- Modiano D, Luoni G, Petrarca V, et al. (2001). "HLA class I in three West African ethnic groups: genetic distances from sub-Saharan and Caucasoid populations". Tissue Antigens. 57 (2): 128–37. doi:10.1034/j.1399-0039.2001.057002128.x. PMID 11260507.
- Torimiro JN, Carr JK, Wolfe ND, et al. (2006). "HLA class I diversity among rural rainforest inhabitants in Cameroon: identification of A*2612-B*4407 haplotype". Tissue Antigens. 67 (1): 30–7. doi:10.1111/j.1399-0039.2005.00527.x. PMID 16451198.
- Sánchez-Velasco P, Karadsheh NS, García-Martín A, Ruíz de Alegría C, Leyva-Cobián F (2001). "Molecular analysis of HLA allelic frequencies and haplotypes in Jordanians and comparison with other related populations". Hum. Immunol. 62 (9): 901–9. doi:10.1016/S0198-8859(01)00289-0. PMID 11543892.