Neutrophil extracellular traps
Neutrophil extracellular traps (NETs) are networks of extracellular fibers, primarily composed of DNA from neutrophils, which bind pathogens.[2] Neutrophils are the immune system's first line of defense against infection and have conventionally been thought to kill invading pathogens through two strategies: engulfment of microbes and secretion of anti-microbials. In 2004, a novel third function was identified: formation of NETs. NETs allow neutrophils to kill extracellular pathogens while minimizing damage to the host cells.[3] Upon in vitro activation with the pharmacological agent phorbol myristate acetate (PMA), Interleukin 8 (IL-8) or lipopolysaccharide (LPS), neutrophils release granule proteins and chromatin to form an extracellular fibril matrix known as NET through an active process.[2]
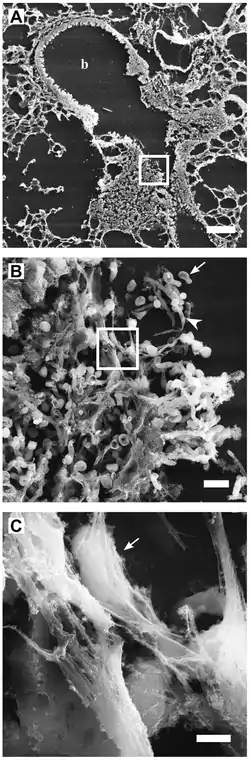
Structure and composition
High-resolution scanning electron microscopy has shown that NETs consist of stretches of DNA and globular protein domains with diameters of 15-17 nm and 25 nm, respectively. These aggregate into larger threads with a diameter of 50 nm.[2] However, under flow conditions, NETs can form much larger structures, reaching hundreds of nanometers in length and width.[4]
Analysis by immunofluorescence corroborated that NETs contain proteins from azurophilic granules (neutrophil elastase, cathepsin G and myeloperoxidase), specific granules (lactoferrin), tertiary granules (gelatinase), and the cytoplasm; however, CD63, actin, tubulin and various other cytoplasmatic proteins are not present in NETs.[2][5]
Anti-microbial activity
NETs disarm pathogens with antimicrobial proteins such as neutrophil elastase, cathepsin G and histones that have a high affinity for DNA.[6] NETs provide for a high local concentration of antimicrobial components and bind, disarm, and kill microbes extracellularly independent of phagocytic uptake. In addition to their antimicrobial properties, NETs may serve as a physical barrier that prevents further spread of the pathogens. Furthermore, delivering the granule proteins into NETs may keep potentially injurious proteins like proteases from diffusing away and inducing damage in tissue adjacent to the site of inflammation.
More recently, it has also been shown that not only bacteria but also pathogenic fungi such as Candida albicans induce neutrophils to form NETs that capture and kill C. albicans hyphal as well as yeast-form cells.[7] NETs have also been documented in association with Plasmodium falciparum infections in children.[8]
While it was originally proposed that NETs would be formed in tissues at a site of bacterial/yeast infection, NETs have also been shown to form within blood vessels during sepsis (specifically in the lung capillaries and liver sinusoids). Intra-vascular NET formation is tightly controlled and is regulated by platelets, which sense severe infection via platelet TLR4 and then bind to and activate neutrophils to form NETs. Platelet-induced NET formation occurs very rapidly (in minutes) and may or may not result in death of the neutrophils.[9] NETs formed in blood vessels can catch circulating bacteria as they pass through the vessels. Trapping of bacteria under flow has been imaged directly in flow chambers in vitro and intravital microscopy demonstrated that bacterial trapping occurs in the liver sinusoids and lung capillaries (sites where platelets bind neutrophils).[4]
NETosis
NET activation and release, or NETosis, is a dynamic process that can come in two forms, suicidal and vital NETosis. Overall, many of the key components of the process are similar for both types of NETosis, however, there are key differences in stimuli, timing, and ultimate end result.[10]
Activation pathway
The full NETosis activation pathway is still under investigation but a few key proteins have been identified and slowly a full picture of the pathway is emerging. The process is thought to begin with NADPH oxidase activation of protein-arginine deiminase 4 (PAD4) via reactive-oxygen species (ROS) intermediaries. PAD4 is responsible for the citrullination of histones in the neutrophil, resulting in decondensation of chromatin. Azurophilic granule proteins such as myeloperoxidase (MPO) and neutrophil elastase (NE) then enter the nucleus and further the decondensation process, resulting in the rupture of the nuclear envelope. The uncondensed chromatin enters the cytoplasm where additional granule and cytoplasmic proteins are added to the early-stage NET. The end result of the process then depends on which NETosis pathway is activated.[10]
Suicidal NETosis
Suicidal NETosis was first described in a 2007 study that noted that the release of NETs resulted in neutrophil death through a different pathway than apoptosis or necrosis.[11] In suicidal NETosis, the intracellular NET formation is followed by the rupture of the plasma membrane, releasing it into the extracellular space. This NETosis pathway can be initiated through activation of Toll-like receptors (TLRs), Fc receptors, and complement receptors with various ligands such as antibodies, PMA, and so on.[10][12] The current understanding is that upon activation of these receptors, downstream signaling results in the release of calcium from the endoplasmic reticulum. This intracellular influx of calcium in turn activates NADPH oxidase, resulting in activation of the NETosis pathway as described above.[12] Of note, suicidal NETosis can take hours, even with high levels of PMA stimulation, while vital NETosis that can be completed in a matter of minutes.[10]
Vital NETosis
Vital NETosis can be stimulated by bacterial lipopolysaccharide (LPS), other "bacterial products, TLR4-activated platelets, or complement proteins in tandem with TLR2 ligands."[10] Vital NETosis is made possible through the blebbing of the nucleus, resulting in a DNA-filled vesicle that is exocytosed and leaves the plasma membrane intact.[10] Its rapid formation and release does not result in neutrophil death, however, the cell is without DNA, raising questions about whether a cell without DNA can be considered alive. It has been noted that neutrophils can continue to phagocytose and kill microbes after vital NETosis, highlighting the neutrophil's anti-microbial versatility.[12]
Regulation
The formation of NETs is regulated by the lipoxygenase pathway – during certain forms of activation (including contact with bacteria) neutrophil 5-lipoxygenase forms 5-HETE-phospholipids that inhibit NET formation.[13] Evidence from laboratory experiments suggests that NETs are cleaned away by macrophages that phagocytose and degrade them.[14]
NET-associated host damage
NETs might also have a deleterious effect on the host, because the extracellular exposure of histone complexes could play a role during the development of autoimmune diseases like systemic lupus erythematosus.[15] NETs could also play a role in inflammatory diseases, as NETs could be identified in preeclampsia, a pregnancy-related inflammatory disorder in which neutrophils are known to be activated.[16] NETs have also been reported in the colon mucosa of patients with the inflammatory bowel disease ulcerative colitis.[17] NETs have also been associated with the production of IgG antinuclear double stranded DNA antibodies in children infected with P. falciparum malaria.[8] NETs have also been found in cancer patients.[18] Preclinical research suggests that NETs are jointly responsible for cancer-related pathologies like thrombosis, organ failure and metastasis formation.[19]
NETs have been shown to contribute to the pathogenesis of HIV/SIV. NETs are capable of capturing HIV virions and destroying them.[20] There is an increase in NET production throughout the course of HIV/SIV, which is reduced by ART. In addition, NETs are able to capture and kill various immune cell groups such as CD4+ and CD8+ T cells, B cells, and monocytes. This effect is seen not only with neutrophils in the blood, but also in various tissues such as the gut, lung, liver, and blood vessels. NETs possibly contribute to the hypercoagulable state in HIV by trapping platelets, and expressing tissue factor.[21]
NETs also have a role in thrombosis and have been associated with stroke.[22][23][24]
These observations suggest that NETs might play an important role in the pathogenesis of infectious, inflammatory and thrombotic disorders.[25][26][27]
Due to the charged and 'sticky' nature of NETs, they may become a problem in Cystic fibrosis sufferers, by increasing sputum viscosity. Treatments have focused on breaking down DNA within sputum, which is largely composed of host NET DNA.
A small study published in the journal JAMA Cardiology suggested that NETs played a major role in COVID-19 patients who developed ST-elevation myocardial infarctions.[28]
References
- Urban, Constantin F.; Ermert, David; Schmid, Monika; Abu-Abed, Ulrike; Goosmann, Christian; Nacken, Wolfgang; Brinkmann, Volker; Jungblut, Peter R.; Zychlinsky, Arturo; Levitz, Stuart M. (30 October 2009). "Neutrophil Extracellular Traps Contain Calprotectin, a Cytosolic Protein Complex Involved in Host Defense against Candida albicans". PLOS Pathogens. 5 (10): e1000639. doi:10.1371/journal.ppat.1000639. PMC 2763347. PMID 19876394.
- Brinkmann, Volker; Ulrike Reichard; Christian Goosmann; Beatrix Fauler; Yvonne Uhlemann; David S. Weiss; Yvette Weinrauch; Arturo Zychlinsky (2004-03-05). "Neutrophil Extracellular Traps Kill Bacteria". Science. 303 (5663): 1532–1535. Bibcode:2004Sci...303.1532B. doi:10.1126/science.1092385. PMID 15001782.
- Nirmala GJ and Lopus M (2020) Cell death mechanisms in eukaryotes. Cell Biol Toxicol, 36, 145–164. doi: /10.1007/s10565-019-09496-2. PMID: 31820165
- Clark SR, Ma AC, Tavener SA, McDonald B, Goodarzi Z, Kelly MM, Patel KD, Chakrabarti S, McAvoy E, Sinclair GD, Keys EM, Allen-Vercoe E, Devinney R, Doig CJ, Green FH, Kubes P (2007). "Platelet Toll-Like Receptor-4 Activates Neutrophil Extracellular Traps to Ensnare Bacteria in Endotoxemic and Septic Blood". Nature Medicine. 13 (4): 463–9. doi:10.1038/nm1565. PMID 17384648.
- Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W, Brinkmann V, Jungblut PR, Zychlinsky A (2009). "Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans". PLOS Pathogens. 5 (10): e1000639. doi:10.1371/journal.ppat.1000639. PMC 2763347. PMID 19876394.
- Thomas MP, Whangbo J, McCrossan G, et al. (June 2014). "Leukocyte protease binding to nucleic acids promotes nuclear localization and cleavage of nucleic acid binding proteins". Journal of Immunology. 192 (11): 5390–7. doi:10.4049/jimmunol.1303296. PMC 4041364. PMID 24771851.
- Urban, CF; Reichard U; Brinkmann V; Zychlinsky A (April 2006). "Neutrophil extracellular traps capture and kill Candida albicans yeast and hyphal forms". Cell Microbiol. 8 (4): 668–76. doi:10.1111/j.1462-5822.2005.00659.x. PMID 16548892.
- Baker VS, Imade GE, Molta NB, Tawde P, Pam SD, Obadofin MO, Sagay SA, Egah DZ, Iya D, Afolabi BB, Baker M, Ford K, Ford R, Roux KH, Keller TC (February 2008). "Cytokine-associated neutrophil extracellular traps and antinuclear antibodies in Plasmodium falciparum infected children under six years of age". Malaria Journal. 7 (41): 41. doi:10.1186/1475-2875-7-41. PMC 2275287. PMID 18312656.
- Caudrillier, Axelle; Kessenbrock, Kai; Gilliss, Brian; Nguyen, John; Marques, Marisa; Monestier, Marc; Toy, Pearl; Werb, Zena; Looney, Mark (2 Jul 2012). "Platelets induce neutrophil extracellular traps in transfusion-related acute lung injury". The Journal of Clinical Investigation. 122 (7): 2661–71. doi:10.1172/JCI61303. PMC 3386815. PMID 22684106.
- Jorch, Selina K.; Kubes, Paul (March 2017). "An emerging role for neutrophil extracellular traps in noninfectious disease". Nature Medicine. 23 (3): 279–287. doi:10.1038/nm.4294. ISSN 1078-8956. PMID 28267716.
- Fuchs, Tobias A.; Abed, Ulrike; Goosmann, Christian; Hurwitz, Robert; Schulze, Ilka; Wahn, Volker; Weinrauch, Yvette; Brinkmann, Volker; Zychlinsky, Arturo (2007-01-15). "Novel cell death program leads to neutrophil extracellular traps". The Journal of Cell Biology. 176 (2): 231–241. doi:10.1083/jcb.200606027. ISSN 0021-9525. PMC 2063942. PMID 17210947.
- Yang, Hang; Biermann, Mona Helena; Brauner, Jan Markus; Liu, Yi; Zhao, Yi; Herrmann, Martin (2016-08-12). "New Insights into Neutrophil Extracellular Traps: Mechanisms of Formation and Role in Inflammation". Frontiers in Immunology. 7: 302. doi:10.3389/fimmu.2016.00302. ISSN 1664-3224. PMC 4981595. PMID 27570525.
- Clark, SR; Guy CJ; Scurr MJ; Taylor PR; Kift-Morgan AP; Hammond VJ; Thomas CP; Coles B; Roberts GW; Eberl M; Jones SA; Topley N; Kotecha S; O'Donnell VB (2011). "Esterified eicosanoids are acutely generated by 5-lipoxygenase in primary human neutrophils and in human and murine infection". Blood. 117 (6): 2033–43. doi:10.1182/blood-2010-04-278887. PMC 3374621. PMID 21177434.
- Farrera, c; Fadeel B (2013). "Macrophage Clearance of Neutrophil Extracellular Traps Is a Silent Process". Journal of Immunology. 191 (5): 2647–56. doi:10.4049/jimmunol.1300436. PMID 23904163.
- Hakkim A, Fürnrohr BG, Amann K, Laube B, Abed UA, Brinkmann V, Herrmann M, Voll RE, Zychlinsky A (2010). "Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis". Proc Natl Acad Sci U S A. 107 (21): 9813–8. Bibcode:2010PNAS..107.9813H. doi:10.1073/pnas.0909927107. PMC 2906830. PMID 20439745.
- Gupta, AK; Hasler P; Holzgreve W; Gebhardt S; Hahn S. (November 2005). "Induction of neutrophil extracellular DNA lattices by placental microparticles and IL-8 and their presence in preeclampsia". Hum Immunol. 66 (11): 1146–54. doi:10.1016/j.humimm.2005.11.003. PMID 16571415.
- Bennike, Tue Bjerg; Carlsen, Thomas Gelsing; Ellingsen, Torkell; Bonderup, Ole Kristian; Glerup, Henning; Bøgsted, Martin; Christiansen, Gunna; Birkelund, Svend; Stensballe, Allan (2015). "Neutrophil Extracellular Traps in Ulcerative Colitis". Inflammatory Bowel Diseases. 21 (9): 2052–2067. doi:10.1097/mib.0000000000000460. PMC 4603666. PMID 25993694.
- Rayes, Roni F.; Mouhanna, Jack G.; Nicolau, Ioana; Bourdeau, France; Giannias, Betty; Rousseau, Simon; Quail, Daniela; Walsh, Logan; Sangwan, Veena; Bertos, Nicholas; Cools-Lartigue, Jonathan (2019-08-22). "Primary tumors induce neutrophil extracellular traps with targetable metastasis-promoting effects". JCI Insight. 4 (16): e128008. doi:10.1172/jci.insight.128008. ISSN 2379-3708. PMC 6777835. PMID 31343990.
- Cedervall, J.; Zhang, Y.; Olsson, A.-K. (2016-08-01). "Tumor-Induced NETosis as a Risk Factor for Metastasis and Organ Failure". Cancer Research. 76 (15): 4311–4315. doi:10.1158/0008-5472.CAN-15-3051. ISSN 0008-5472. PMID 27402078.
- Saitoh, Tatsuya; Komano, Jun; Saitoh, Yasunori; Misawa, Takuma; Takahama, Michihiro; Kozaki, Tatsuya; Uehata, Takuya; Iwasaki, Hidenori; Omori, Hiroko (July 2012). "Neutrophil Extracellular Traps Mediate a Host Defense Response to Human Immunodeficiency Virus-1". Cell Host & Microbe. 12 (1): 109–116. doi:10.1016/j.chom.2012.05.015. ISSN 1931-3128. PMID 22817992.
- Sivanandham, Ranjit; Brocca-Cofano, Egidio; Krampe, Noah; Falwell, Elizabeth; Kilapandal Venkatraman, Sindhuja Murali; Ribeiro, Ruy M.; Apetrei, Cristian; Pandrea, Ivona (2018-09-11). "Neutrophil extracellular trap production contributes to pathogenesis in SIV-infected nonhuman primates". Journal of Clinical Investigation. 128 (11): 5178–5183. doi:10.1172/jci99420. ISSN 1558-8238. PMC 6205390. PMID 30204591.
- Laridan, Elodie; Denorme, Frederik; Desender, Linda; François, Olivier; Andersson, Tommy; Deckmyn, Hans; Vanhoorelbeke, Karen; De Meyer, Simon F. (August 2, 2017). "Neutrophil extracellular traps in ischemic stroke thrombi". Annals of Neurology. 82 (2): 223–232. doi:10.1002/ana.24993. PMID 28696508.
- Ducroux, Celina; Di Meglio, Lucas; Loyau, Stephane; Delbosc, Sandrine; Boisseau, William; Deschildre, Catherine; Ben Maacha, Malek; Blanc, Raphael; Redjem, Hocine; Ciccio, Gabriele; Smajda, Stanislas; Fahed, Robert; Michel, Jean-Baptiste; Piotin, Michel; Salomon, Laurence; Mazighi, Mikael; Ho-Tin-Noe, Benoit; Desilles, Jean-Philippe (March 2, 2018). "Thrombus Neutrophil Extracellular Traps Content Impair tPA-Induced Thrombolysis in Acute Ischemic Stroke". Stroke. 49 (3): 754–757. doi:10.1161/STROKEAHA.117.019896. PMID 29438080.
- Vallés, Juana; Lago, Aída; Santos, María Teresa; Latorre, Ana María; Tembl, José I.; Salom, Juan B.; Nieves, Candela; Moscardó, Antonio (October 5, 2017). "Neutrophil extracellular traps are increased in patients with acute ischemic stroke: prognostic significance". Thrombosis and Haemostasis. 117 (10): 1919–1929. doi:10.1160/TH17-02-0130. PMID 28837206.
- Fuchs TA, Brill A, Duerschmied D, Schatzberg D, Monestier M, Myers DD, Wrobleski SK, Wakefield TW, Hartwig JH, Wagner DD (Sep 7, 2010). "Extracellular DNA traps promote thrombosis". Proc Natl Acad Sci U S A. 107 (36): 15880–5. Bibcode:2010PNAS..10715880F. doi:10.1073/pnas.1005743107. PMC 2936604. PMID 20798043.
- Brill A, Fuchs TA, Savchenko AS, Thomas GM, Martinod K, De Meyer SF, Bhandari AA, Wagner DD (Nov 1, 2011). "Neutrophil Extracellular Traps Promote Deep Vein Thrombosis in Mice". Journal of Thrombosis and Haemostasis. 10 (1): 136–144. doi:10.1111/j.1538-7836.2011.04544.x. PMC 3319651. PMID 22044575.
- Borissoff, JI; ten Cate, H (September 2011). "From neutrophil extracellular traps release to thrombosis: an overshooting host-defense mechanism?". Journal of Thrombosis and Haemostasis. 9 (9): 1791–4. doi:10.1111/j.1538-7836.2011.04425.x. PMID 21718435.
- Blasco, Ana; Coronado, María-José; Hernández-Terciado, Fernando; Martín, Paloma; Royuela, Ana; Ramil, Elvira; García, Diego; Goicolea, Javier; Del Trigo, María; Ortega, Javier; Escudier, Juan M. (2020-12-29). "Assessment of Neutrophil Extracellular Traps in Coronary Thrombus of a Case Series of Patients With COVID-19 and Myocardial Infarction". JAMA Cardiology. doi:10.1001/jamacardio.2020.7308. ISSN 2380-6583.
External links
- Frontiers in Molecular Innate Immunity research topic about NET
- "Molecular mechanisms involved in neutrophil extracellular trap (NET) formation", PhD thesis, Jyaysi Desai, Ludwig Maximilian University of Munich, Germany. Chemistry winner, Dance Your PhD, 2015.