Lipopolysaccharide
Lipopolysaccharides (LPS), also known as endotoxins, are large molecules consisting of a lipid and a polysaccharide composed of O-antigen, outer core and inner core joined by a covalent bond; they are found in the outer membrane of Gram-negative bacteria.

The term lipooligosaccharide ("LOS") is used to refer to a low-molecular-weight form of bacterial lipopolysaccharides.
Discovery
The toxic activity of LPS was first discovered and termed "endotoxin" by Richard Friedrich Johannes Pfeiffer, who distinguished between exotoxins, which he classified as a toxin that is released by bacteria into the surrounding environment, and endotoxins, which he considered to be a toxin kept "within" the bacterial cell and released only after destruction of the bacterial cell wall.[1]:84 Subsequent work showed that release of LPS from gram negative microbes does not necessarily require the destruction of the bacterial cell wall, but rather, LPS is secreted as part of the normal physiological activity of membrane vesicle trafficking in the form of bacterial outer membrane vesicles (OMVs), which may also contain other virulence factors and proteins.[2]
Today, the term 'endotoxin' is mostly used synonymously with LPS,[3] although there are a few endotoxins that are not related to LPS, such as the so-called delta endotoxin proteins secreted by Bacillus thuringiensis.
Functions in bacteria
LPS is the major component of the outer membrane of Gram-negative bacteria, contributing greatly to the structural integrity of the bacteria, and protecting the membrane from certain kinds of chemical attack. LPS also increases the negative charge of the cell membrane and helps stabilize the overall membrane structure. It is of crucial importance to many Gram-negative bacteria, which die if it is mutated or removed; however, it appears that LPS is nonessential in at least some Gram-negative bacteria, such as Neisseria meningitidis, Moraxella catarrhalis, and Acinetobacter baumannii.[4] LPS induces a strong response from normal animal immune systems. It has also been implicated in non-pathogenic aspects of bacterial ecology, including surface adhesion, bacteriophage sensitivity, and interactions with predators such as amoebae.
LPS is required for the proper conformation of omptin activity; however, smooth LPS will sterically hinder omptins.
Composition
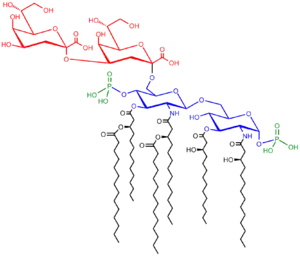
It comprises three parts:
- O antigen (or O polysaccharide)
- Core oligosaccharide
- Lipid A
O-antigen
A repetitive glycan polymer contained within an LPS is referred to as the O antigen, O polysaccharide, or O side-chain of the bacteria. The O antigen is attached to the core oligosaccharide, and comprises the outermost domain of the LPS molecule. The composition of the O chain varies from strain to strain. For example, there are over 160 different O antigen structures produced by different E. coli strains.[5] The presence or absence of O chains determines whether the LPS is considered rough or smooth. Full-length O-chains would render the LPS smooth, whereas the absence or reduction of O-chains would make the LPS rough.[6] Bacteria with rough LPS usually have more penetrable cell membranes to hydrophobic antibiotics, since a rough LPS is more hydrophobic.[7] O antigen is exposed on the very outer surface of the bacterial cell, and, as a consequence, is a target for recognition by host antibodies.
Core
The core domain always contains an oligosaccharide component that attaches directly to lipid A and commonly contains sugars such as heptose and 3-Deoxy-D-manno-oct-2-ulosonic acid (also known as KDO, keto-deoxyoctulosonate).[8] The LPS cores of many bacteria also contain non-carbohydrate components, such as phosphate, amino acids, and ethanolamine substituents.
Lipid A
Lipid A is, in normal circumstances, a phosphorylated glucosamine disaccharide decorated with multiple fatty acids. These hydrophobic fatty acid chains anchor the LPS into the bacterial membrane, and the rest of the LPS projects from the cell surface. The lipid A domain is responsible for much of the toxicity of Gram-negative bacteria. When bacterial cells are lysed by the immune system, fragments of membrane containing lipid A are released into the circulation, causing fever, diarrhea, and possible fatal endotoxic shock (also called septic shock). The Lipid A moiety is a very conserved component of the LPS.[9] However Lipid A structure varies among bacterial species and Lipid A structure defines an overall host immune activation.[10]
Lipooligosaccharides
Lipooligosaccharides (LOS) are glycolipids found in the outer membrane of some types of Gram-negative bacteria, such as Neisseria spp. and Haemophilus spp. The term is synonymous with the low molecular weight form of bacterial LPS.[11] LOS plays a central role in maintaining the integrity and functionality of the outer membrane of the Gram negative cell envelope. Lipooligosaccharides play an important role in the pathogenesis of certain bacterial infections because they are capable of acting as immunostimulators and immunomodulators.[11] Furthermore, LOS molecules are responsible for the ability of some bacterial strains to display molecular mimicry and antigenic diversity, aiding in the evasion of host immune defenses and thus contributing to the virulence of these bacterial strains.
Chemically, lipooligosaccharides lack O-antigens and possess only a lipid A-based outer membrane-anchoring moiety, and an oligosaccharide core.[12] In the case of Neisseria meningitidis, the lipid A portion of the molecule has a symmetrical structure and the inner core is composed of 3-deoxy-D-manno-2-octulosonic acid (KDO) and heptose (Hep) moieties. The outer core oligosaccharide chain varies depending on the bacterial strain.[11][12] The term lipooligosaccharide is used to refer to the low molecular weight form of bacterial lipopolysaccharides, which can be categorized into two forms: the high molecular weight (Mr, or smooth) form possesses a high molecular weight, repeating polysaccharide O-chain, while the low molecular weight (low-Mr or rough) form, lacks the O-chain but possesses a short oligosaccharide in its place.[11]
LPS modifications
The making of LPS can be modified in order to present a specific sugar structure. Those can be recognised by either other LPS (which enables to inhibit LPS toxins) or glycosyltransferases that use those sugar structure to add more specific sugars. A highly conserved host enzyme may detoxify LPS when it enters, or is produced in, animal tissues. It may also convert LPS in the intestine into an LPS inhibitor. Neutrophils, macrophages and dendritic cells produce this lipase, acyloxyacyl hydrolase (AOAH), which inactivates LPS by removing the two secondary acyl chains from lipid A to produce tetraacyl LPS. If they are given LPS parenterally, mice that lack AOAH develop high titers of non-specific antibodies, develop prolonged hepatomegaly, and experience prolonged endotoxin tolerance. LPS inactivation may be required for animals to restore homeostasis after parenteral LPS exposure.[13] Although mice have many other mechanisms for inhibiting LPS signaling, none is able to prevent these changes in animals that lack AOAH.
Biosynthesis and transport
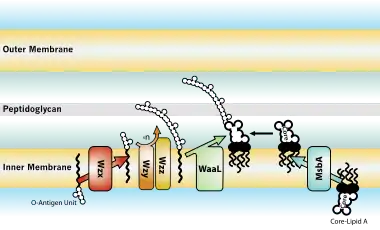
Biological effects on hosts infected with Gram-negative bacteria
Immune response
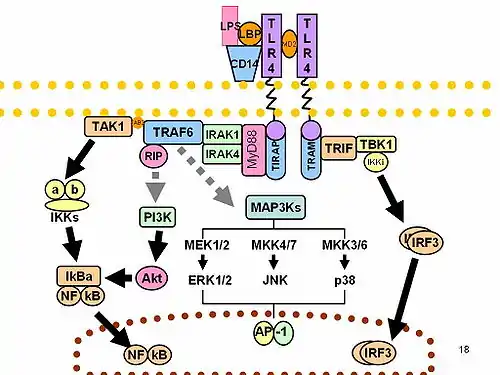
LPS acts as the prototypical endotoxin because it binds the CD14/TLR4/MD2 receptor complex in many cell types, but especially in monocytes, dendritic cells, macrophages and B cells, which promotes the secretion of pro-inflammatory cytokines, nitric oxide, and eicosanoids.[16]
As part of the cellular stress response, superoxide is one of the major reactive oxygen species induced by LPS in various cell types that express TLR (toll-like receptor).
LPS is also an exogenous pyrogen (fever-inducing substance).
Being of crucial importance to Gram-negative bacteria, these molecules make candidate targets for new antimicrobial agents.
Some researchers doubt reports of generalized toxic effects attributed to all lipopolysaccharides, in particular, for cyanobacteria.[17]
LPS function has been under experimental research for several years due to its role in activating many transcription factors. LPS also produces many types of mediators involved in septic shock. Humans are much more sensitive to LPS than other animals (e.g., mice). A dose of 1 µg/kg induces shock in humans, but mice will tolerate a dose up to a thousand times higher.[18] This may relate to differences in the level of circulating natural antibodies between the two species.[19][20] Said et al. showed that LPS causes an IL-10-dependent inhibition of CD4 T-cell expansion and function by up-regulating PD-1 levels on monocytes which leads to IL-10 production by monocytes after binding of PD-1 by PD-L1.[21]
Endotoxins are in large part responsible for the dramatic clinical manifestations of infections with pathogenic Gram-negative bacteria, such as Neisseria meningitidis, the pathogens that causes meningococcal disease, including meningococcemia, Waterhouse–Friderichsen syndrome, and meningitis. Alkaline phosphatase prevents intestinal inflammation (and "leaky gut") from bacteria by dephosphorylating the Lipid A portion of LPS.[22][23][24]
Bruce Beutler was awarded a portion of the 2011 Nobel Prize in Physiology or Medicine for his work demonstrating that TLR4 is the LPS receptor.[25][26]
Portions of the LPS from several bacterial strains have been shown to be chemically similar to human host cell surface molecules; the ability of some bacteria to present molecules on their surface which are chemically identical or similar to the surface molecules of some types of host cells is termed molecular mimicry.[27] For example, in Neisseria meningitidis L2,3,5,7,9, the terminal tetrasaccharide portion of the oligosaccharide (lacto-N-neotetraose) is the same tetrasaccharide as that found in paragloboside, a precursor for ABH glycolipid antigens found on human erythrocytes.[11] In another example, the terminal trisaccharide portion (lactotriaose) of the oligosaccharide from pathogenic Neisseria spp. LOS is also found in lactoneoseries glycosphingolipids from human cells.[11] Most meningococci from groups B and C, as well as gonococci, have been shown to have this trisaccharide as part of their LOS structure.[11] The presence of these human cell surface 'mimics' may, in addition to acting as a 'camouflage' from the immune system, play a role in the abolishment of immune tolerance when infecting hosts with certain human leukocyte antigen (HLA) genotypes, such as HLA-B35.[11]
Recently, a new study published has discovered that LPS can be sensed directly by hematopoietic stem cells (HSCs) through the bonding with TLR4, causing them to proliferate in reaction to a systemic infection. This response activate the TLR4-TRIF-ROS-p38 signaling within the HSCs and through a sustained TLR4 activation can cause a proliferative stress, leading to impair their competitive repopulating ability.[28] Infection in mice using S. typhimurium showed similar results, validating the experimental model also in vivo.
Effect of variability on immune response
O-antigens (the outer carbohydrates) are the most variable portion of the LPS molecule, imparting the antigenic specificity. In contrast, lipid A is the most conserved part. However, lipid A composition also may vary (e.g., in number and nature of acyl chains even within or between genera). Some of these variations may impart antagonistic properties to these LPS. For example, Rhodobacter sphaeroides diphosphoryl lipid A (RsDPLA) is a potent antagonist of LPS in human cells, but is an agonist in hamster and equine cells.
It has been speculated that conical Lipid A (e.g., from E. coli) are more agonistic, less conical lipid A like those of Porphyromonas gingivalis may activate a different signal (TLR2 instead of TLR4), and completely cylindrical lipid A like that of Rhodobacter sphaeroides is antagonistic to TLRs.[29][30]
LPS gene clusters are highly variable between different strains, subspecies, species of bacterial pathogens of plants and animals.[31][32]
Normal human blood serum contains anti-LOS antibodies that are bactericidal and patients that have infections caused by serotypically distinct strains possess anti-LOS antibodies that differ in their specificity compared with normal serum.[33] These differences in humoral immune response to different LOS types can be attributed to the structure of the LOS molecule, primarily within the structure of the oligosaccharide portion of the LOS molecule.[33] In Neisseria gonorrhoeae it has been demonstrated that the antigenicity of LOS molecules can change during an infection due to the ability of these bacteria to synthesize more than one type of LOS,[33] a characteristic known as phase variation. Additionally, Neisseria gonorrhoeae, as well as Neisseria meningitidis and Haemophilus influenzae,[11] are capable of further modifying their LOS in vitro, for example through sialylation (modification with sialic acid residues), and as a result are able to increase their resistance to complement-mediated killing [33] or even down-regulate complement activation[11] or evade the effects of bactericidal antibodies.[11] Sialylation may also contribute to hindered neutrophil attachment and phagocytosis by immune system cells as well as a reduced oxidative burst.[11] Haemophilus somnus, a pathogen of cattle, has also been shown to display LOS phase variation, a characteristic which may help in the evasion of bovine host immune defenses.[34] Taken together, these observations suggest that variations in bacterial surface molecules such as LOS can help the pathogen evade both the humoral (antibody and complement-mediated) and the cell-mediated (killing by neutrophils, for example) host immune defenses.
Non-canonical pathways of LPS recognition
Recently, it was shown that in addition to TLR4 mediated pathways, certain members of the family of the transient receptor potential ion channels recognize LPS.[35] LPS-mediated activation of TRPA1 was shown in mice[36] and Drosophila melanogaster flies.[37] At higher concentrations, LPS activates other members of the sensory TRP channel family as well, such as TRPV1, TRPM3 and to some extent TRPM8.[38] LPS is recognized by TRPV4 on epithelial cells. TRPV4 activation by LPS was necessary and sufficient to induce nitric oxide production with a bactericidal effect.[39]
Health effects
Endotoxemia
The presence of endotoxins in the blood is called endotoxemia. It can lead to septic shock, if the immune response is severely pronounced.[40]
Moreover, endotoxemia of intestinal origin, especially, at the host-pathogen interface, is considered to be an important factor in the development of alcoholic hepatitis,[41] which is likely to develop on the basis of the small bowel bacterial overgrowth syndrome and an increased intestinal permeability.[42]
Lipid A may cause uncontrolled activation of mammalian immune systems with production of inflammatory mediators that may lead to septic shock.[12] This inflammatory reaction is mediated by Toll-like receptor 4 which is responsible for immune system cell activation.[12] Damage to the endothelial layer of blood vessels caused by these inflammatory mediators can lead to capillary leak syndrome, dilation of blood vessels and a decrease in cardiac function and can lead to septic shock.[43] Pronounced complement activation can also be observed later in the course as the bacteria multiply in the blood.[43] High bacterial proliferation triggering destructive endothelial damage can also lead to disseminated intravascular coagulation (DIC) with loss of function of certain internal organs such as the kidneys, adrenal glands and lungs due to compromised blood supply. The skin can show the effects of vascular damage often coupled with depletion of coagulation factors in the form of petechiae, purpura and ecchymoses. The limbs can also be affected, sometimes with devastating consequences such as the development of gangrene, requiring subsequent amputation.[43] Loss of function of the adrenal glands can cause adrenal insufficiency and additional hemorrhage into the adrenals causes Waterhouse-Friderichsen syndrome, both of which can be life-threatening. It has also been reported that gonococcal LOS can cause damage to human fallopian tubes.[33]
Auto-immune disease
The molecular mimicry of some LOS molecules is thought to cause autoimmune-based host responses, such as flareups of multiple sclerosis.[11][27] Other examples of bacterial mimicry of host structures via LOS are found with the bacteria Helicobacter pylori and Campylobacter jejuni, organisms which cause gastrointestinal disease in humans, and Haemophilus ducreyi which causes chancroid. Certain C. jejuni LPS serotypes (attributed to certain tetra- and pentasaccharide moieties of the core oligosaccharide) have also been implicated with Guillain–Barré syndrome and a variant of Guillain–Barré called Miller-Fisher syndrome.[11]
Link to obesity
Epidemiological studies have shown that increased endotoxin load, which can be a result of increased populations of endotoxin-producing bacteria in the intestinal tract, is associated with certain obesity-related patient groups.[44][45][46] Other studies have shown that purified endotoxin from Escherichia coli can induce obesity and insulin-resistance when injected into germ-free mouse models.[47] A more recent study has uncovered a potentially contributing role for Enterobacter cloacae B29 toward obesity and insulin resistance in a human patient.[48] The presumed mechanism for the association of endotoxin with obesity is that endotoxin induces an inflammation-mediated pathway accounting for the observed obesity and insulin resistance.[47] Bacterial genera associated with endotoxin-related obesity effects include Escherichia and Enterobacter.
Laboratory research and biotechnology production systems
Lipopolysaccharides are frequent contaminants in plasmid DNA prepared from bacteria or proteins expressed from bacteria, and must be removed from the DNA or protein to avoid contaminating experiments and to avoid toxicity of products manufactured using industrial fermentation.[49]
Also, ovalbumin is frequently contaminated with endotoxins. Ovalbumin is one of the extensively studied proteins in animal models and also an established model allergen for airway hyper-responsiveness (AHR). Commercially available ovalbumin that is contaminated with LPS can fully activate endothelial cells in an in-vitro assay of the first step of inflammation, and it falsifies research results, as it does not accurately reflect the effect of sole protein antigen on animal physiology.
In pharmaceutical production, it is necessary to remove all traces of endotoxin from drug product containers, as even small amounts of endotoxin will cause illness in humans. A depyrogenation oven is used for this purpose. Temperatures in excess of 300 °C are required to break down this substance. A defined endotoxin reduction rate is a correlation between time and temperature. Based on primary packaging material as syringes or vials, a glass temperature of 250 °C and a holding time of 30 minutes is typical to achieve a reduction of endotoxin levels by a factor of 1000.[50]
The standard assay for detecting presence of endotoxin is the Limulus Amebocyte Lysate (LAL) assay, utilizing blood from the Horseshoe crab (Limulus polyphemus).[51] Very low levels of LPS can cause coagulation of the limulus lysate due to a powerful amplification through an enzymatic cascade. However, due to the dwindling population of horseshoe crabs, and the fact that there are factors that interfere with the LAL assay, efforts have been made to develop alternative assays, with the most promising ones being ELISA tests using a recombinant version of a protein in the LAL assay, Factor C.[52]
See also
References
- Parija SC (1 January 2009). Textbook of Microbiology & Immunology. India: Elsevier. ISBN 978-8131221631.
- Kulp A, Kuehn MJ (2010). "Biological functions and biogenesis of secreted bacterial outer membrane vesicles". Annu. Rev. Microbiol. 64: 163–84. doi:10.1146/annurev.micro.091208.073413. PMC 3525469. PMID 20825345.
- Rietschel ET, Kirikae T, Schade FU, Mamat U, Schmidt G, Loppnow H, Ulmer AJ, Zähringer U, Seydel U, Di Padova F (1994). "Bacterial endotoxin: molecular relationships of structure to activity and function". FASEB J. 8 (2): 217–25. doi:10.1096/fasebj.8.2.8119492. PMID 8119492.
- Zhang G, Meredith TC, Kahne D (2013). "On the essentiality of lipopolysaccharide to Gram-negative bacteria". Curr. Opin. Microbiol. 16 (6): 779–785. doi:10.1016/j.mib.2013.09.007. PMC 3974409. PMID 24148302.
- Raetz CR, Whitfield C (2002). "Lipopolysaccharide endotoxins". Annu. Rev. Biochem. 71: 635–700. doi:10.1146/annurev.biochem.71.110601.135414. PMC 2569852. PMID 12045108.
- Rittig MG, Kaufmann A, Robins A, Shaw B, Sprenger H, Gemsa D, Foulongne V, Rouot B, Dornand J (2003). "Smooth and rough lipopolysaccharide phenotypes of Brucella induce different intracellular trafficking and cytokine/chemokine release in human monocytes". J. Leukoc. Biol. 74 (6): 1045–55. doi:10.1189/jlb.0103015. PMID 12960272.
- Tsujimoto H, Gotoh N, Nishino T (1999). "Diffusion of macrolide antibiotics through the outer membrane of Moraxella catarrhalis". J. Infect. Chemother. 5 (4): 196–200. doi:10.1007/s101560050034. PMID 11810516.
- Hershberger C, Binkley SB (1968). "Chemistry and metabolism of 3-deoxy-D-mannooctulosonic acid. I. Stereochemical determination". J. Biol. Chem. 243 (7): 1578–84. PMID 4296687.
- Tzeng YL, Datta A, Kolli VK, Carlson RW, Stephens DS (May 2002). "Endotoxin of Neisseria meningitidis composed only of intact lipid A: inactivation of the meningococcal 3-deoxy-D-manno-octulosonic acid transferase". J. Bacteriol. 184 (9): 2379–88. doi:10.1128/JB.184.9.2379-2388.2002. PMC 134985. PMID 11948150.
- Khan, Mohd M.; Ernst, Orna; Sun, Jing; Fraser, Iain D. C.; Ernst, Robert K.; Goodlett, David R.; Nita-Lazar, Aleksandra (24 June 2018). "Mass Spectrometry-based Structural Analysis and Systems Immunoproteomics Strategies for Deciphering the Host Response to Endotoxin". Journal of Molecular Biology. 430 (17): 2641–2660. doi:10.1016/j.jmb.2018.06.032. ISSN 1089-8638. PMID 29949751.
- Moran AP, Prendergast MM, Appelmelk BJ (1996). "Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease". FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391. Archived from the original on 23 September 2016.
- Kilár A, Dörnyei Á, Kocsis B (2013). "Structural characterization of bacterial lipopolysaccharides with mass spectrometry and on- and off-line separation techniques". Mass Spectrom Rev. 32 (2): 90–117. Bibcode:2013MSRv...32...90K. doi:10.1002/mas.21352. PMID 23165926.
- Munford R, Lu M, Varley AW (2009). Kill the Bacteria...and Also Their Messengers?. Advances in Immunology. 103. pp. 29–48. doi:10.1016/S0065-2776(09)03002-8. ISBN 9780123748324. PMC 2812913. PMID 19755182.
- Wang X, Quinn PJ (2010). "Lipopolysaccharide: Biosynthetic pathway and structure modification". Prog. Lipid Res. 49 (2): 97–107. doi:10.1016/j.plipres.2009.06.002. PMID 19815028.
- Ruiz N, Kahne D, Silhavy TJ (2009). "Transport of lipopolysaccharide across the cell envelope: the long road of discovery". Nat. Rev. Microbiol. 7 (9): 677–83. doi:10.1038/nrmicro2184. PMC 2790178. PMID 19633680.
- Abbas, Abul (2006). Basic Immunology. Elsevier. ISBN 978-1-4160-2974-8.
- Stewart I, Schluter PJ, Shaw GR (2006). "Cyanobacterial lipopolysaccharides and human health - a review". Environ Health. 5: 7. doi:10.1186/1476-069X-5-7. PMC 1489932. PMID 16563160.
- Warren HS, Fitting C, Hoff E, Adib-Conquy M, Beasley-Topliffe L, Tesini B, Liang X, Valentine C, Hellman J, Hayden D, Cavaillon JM (2010). "Resilience to bacterial infection: difference between species could be due to proteins in serum". J. Infect. Dis. 201 (2): 223–32. doi:10.1086/649557. PMC 2798011. PMID 20001600.
- Reid RR, Prodeus AP, Khan W, Hsu T, Rosen FS, Carroll MC (1997). "Endotoxin shock in antibody-deficient mice: unraveling the role of natural antibody and complement in the clearance of lipopolysaccharide". J. Immunol. 159 (2): 970–5. PMID 9218618.
- Boes M, Prodeus AP, Schmidt T, Carroll MC, Chen J (1998). "A critical role of natural immunoglobulin M in immediate defense against systemic bacterial infection". J. Exp. Med. 188 (12): 2381–6. doi:10.1084/jem.188.12.2381. PMC 2212438. PMID 9858525.
- Said EA, Dupuy FP, Trautmann L, Zhang Y, Shi Y, El-Far M, Hill BJ, Noto A, Ancuta P, Peretz Y, Fonseca SG, Van Grevenynghe J, Boulassel MR, Bruneau J, Shoukry NH, Routy JP, Douek DC, Haddad EK, Sekaly RP (2010). "Programmed death-1-induced interleukin-10 production by monocytes impairs CD4+ T cell activation during HIV infection". Nat. Med. 16 (4): 452–9. doi:10.1038/nm.2106. PMC 4229134. PMID 20208540.
- Bates JM, Akerlund J, Mittge E, Guillemin K (2007). "Intestinal alkaline phosphatase detoxifies lipopolysaccharide and prevents inflammation in zebrafish in response to the gut microbiota". Cell Host & Microbe. 2 (6): 371–382. doi:10.1016/j.chom.2007.10.010. PMC 2730374. PMID 18078689.
- Alam SN, Yammine H, Moaven O, Ahmed R, Moss AK, Biswas B, Muhammad N, Biswas R, Raychowdhury A, Kaliannan K, Ghosh S, Ray M, Hamarneh SR, Barua S, Malo NS, Bhan AK, Malo MS, Hodin RA (April 2014). "Intestinal alkaline phosphatase prevents antibiotic-induced susceptibility to enteric pathogens". Annals of Surgery. 259 (4): 715–22. doi:10.1097/sla.0b013e31828fae14. PMC 3855644. PMID 23598380.
- Lallès JP (February 2014). "Intestinal alkaline phosphatase: novel functions and protective effects". Nutrition Reviews. 72 (2): 82–94. doi:10.1111/nure.12082. PMID 24506153.
- Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C, Du X, Birdwell D, Alejos E, Silva M, Galanos C, Freudenberg M, Ricciardi-Castagnoli P, Layton B, Beutler B (1998). "Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene". Science. 282 (5396): 2085–8. Bibcode:1998Sci...282.2085P. doi:10.1126/science.282.5396.2085. PMID 9851930.
- "The 2011 Nobel Prize in Physiology or Medicine - Press Release". www.nobelprize.org. Archived from the original on 23 March 2018. Retrieved 28 April 2018.
- Chastain EM, Miller SD (2012). "Molecular mimicry as an inducing trigger for CNS autoimmune demyelinating disease". Immunol. Rev. 245 (1): 227–38. doi:10.1111/j.1600-065X.2011.01076.x. PMC 3586283. PMID 22168423.
- Takizawa, Hitoshi; Fritsch, Kristin; Kovtonyuk, Larisa V.; Saito, Yasuyuki; Yakkala, Chakradhar; Jacobs, Kurt; Ahuja, Akshay K.; Lopes, Massimo; Hausmann, Annika (3 August 2017). "Pathogen-Induced TLR4-TRIF Innate Immune Signaling in Hematopoietic Stem Cells Promotes Proliferation but Reduces Competitive Fitness". Cell Stem Cell. 21 (2): 225–240.e5. doi:10.1016/j.stem.2017.06.013. ISSN 1875-9777. PMID 28736216.
- Netea MG, van Deuren M, Kullberg BJ, Cavaillon JM, Van der Meer JW (2002). "Does the shape of lipid A determine the interaction of LPS with Toll-like receptors?". Trends Immunol. 23 (3): 135–9. doi:10.1016/S1471-4906(01)02169-X. PMID 11864841.
- Seydel U, Oikawa M, Fukase K, Kusumoto S, Brandenburg K (2000). "Intrinsic conformation of lipid A is responsible for agonistic and antagonistic activity". Eur. J. Biochem. 267 (10): 3032–9. doi:10.1046/j.1432-1033.2000.01326.x. PMID 10806403.
- Reeves PP, Wang L (2002). Genomic organization of LPS-specific loci. Curr. Top. Microbiol. Immunol. Current Topics in Microbiology and Immunology. 264. pp. 109–35. doi:10.1007/978-3-642-56031-6_7. ISBN 978-3-540-42682-0. PMID 12014174.
- Patil PB, Sonti RV (2004). "Variation suggestive of horizontal gene transfer at a lipopolysaccharide (lps) biosynthetic locus in Xanthomonas oryzae pv. oryzae, the bacterial leaf blight pathogen of rice". BMC Microbiol. 4: 40. doi:10.1186/1471-2180-4-40. PMC 524487. PMID 15473911.
- Yamasaki R, Kerwood DE, Schneider H, Quinn KP, Griffiss JM, Mandrell RE (1994). "The structure of lipooligosaccharide produced by Neisseria gonorrhoeae, strain 15253, isolated from a patient with disseminated infection. Evidence for a new glycosylation pathway of the gonococcal lipooligosaccharide". J. Biol. Chem. 269 (48): 30345–51. PMID 7982947.
- Howard MD, Cox AD, Weiser JN, Schurig GG, Inzana TJ (2000). "Antigenic diversity of Haemophilus somnus lipooligosaccharide: phase-variable accessibility of the phosphorylcholine epitope". J. Clin. Microbiol. 38 (12): 4412–9. doi:10.1128/JCM.38.12.4412-4419.2000. PMC 87614. PMID 11101573.
- Boonen, Brett; Alpizar, Yeranddy; Meseguer, Victor; Talavera, Karel; Boonen, Brett; Alpizar, Yeranddy A.; Meseguer, Victor M.; Talavera, Karel (11 August 2018). "TRP Channels as Sensors of Bacterial Endotoxins". Toxins. 10 (8): 326. doi:10.3390/toxins10080326. PMC 6115757. PMID 30103489.
- Meseguer, Victor; Alpizar, Yeranddy A.; Luis, Enoch; Tajada, Sendoa; Denlinger, Bristol; Fajardo, Otto; Manenschijn, Jan-Albert; Fernández-Peña, Carlos; Talavera, Arturo; Kichko, Tatiana; Navia, Belén; Sánchez, Alicia; Señarís, Rosa; Reeh, Peter; Pérez-García, María Teresa; López-López, José Ramón; Voets, Thomas; Belmonte, Carlos; Talavera, Karel; Viana, Félix (20 January 2014). "TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins". Nature Communications. 5: 3125. Bibcode:2014NatCo...5.3125M. doi:10.1038/ncomms4125. PMC 3905718. PMID 24445575.
- Soldano, Alessia; Alpizar, Yeranddy A; Boonen, Brett; Franco, Luis; López-Requena, Alejandro; Liu, Guangda; Mora, Natalia; Yaksi, Emre; Voets, Thomas; Vennekens, Rudi; Hassan, Bassem A; Talavera, Karel (14 June 2016). "Gustatory-mediated avoidance of bacterial lipopolysaccharides via TRPA1 activation in Drosophila". eLife. 5. doi:10.7554/eLife.13133. PMC 4907694. PMID 27296646.
- Boonen, Brett; Alpizar, Yeranddy A.; Sanchez, Alicia; López-Requena, Alejandro; Voets, Thomas; Talavera, Karel (July 2018). "Differential effects of lipopolysaccharide on mouse sensory TRP channels". Cell Calcium. 73: 72–81. doi:10.1016/j.ceca.2018.04.004. PMID 29689522.
- Alpizar, Yeranddy A.; Boonen, Brett; Sanchez, Alicia; Jung, Carole; López-Requena, Alejandro; Naert, Robbe; Steelant, Brecht; Luyts, Katrien; Plata, Cristina; De Vooght, Vanessa; Vanoirbeek, Jeroen A. J.; Meseguer, Victor M.; Voets, Thomas; Alvarez, Julio L.; Hellings, Peter W.; Hoet, Peter H. M.; Nemery, Benoit; Valverde, Miguel A.; Talavera, Karel (20 October 2017). "TRPV4 activation triggers protective responses to bacterial lipopolysaccharides in airway epithelial cells". Nature Communications. 8 (1): 1059. Bibcode:2017NatCo...8.1059A. doi:10.1038/s41467-017-01201-3. PMC 5651912. PMID 29057902.
- Opal SM (2010). Endotoxins and other sepsis triggers. Contrib Nephrol. Contributions to Nephrology. 167. pp. 14–24. doi:10.1159/000315915. ISBN 978-3-8055-9484-4. PMID 20519895.
- Ceccanti M, Attili A, Balducci G, Attilia F, Giacomelli S, Rotondo C, Sasso GF, Xirouchakis E, Attilia ML (2006). "Acute alcoholic hepatitis". J. Clin. Gastroenterol. 40 (9): 833–41. doi:10.1097/01.mcg.0000225570.04773.5d. PMID 17016141.
- Parlesak A, Schäfer C, Schütz T, Bode JC, Bode C (2000). "Increased intestinal permeability to macromolecules and endotoxemia in patients with chronic alcohol abuse in different stages of alcohol-induced liver disease". J. Hepatol. 32 (5): 742–7. doi:10.1016/S0168-8278(00)80242-1. PMID 10845660.
- Stephens DS, Greenwood B, Brandtzaeg P (2007). "Epidemic meningitis, meningococcaemia, and Neisseria meningitidis". Lancet. 369 (9580): 2196–210. doi:10.1016/S0140-6736(07)61016-2. PMID 17604802.
- Moreno-Navarrete JM, Ortega F, Serino M, Luche E, Waget A, Pardo G, Salvador J, Ricart W, Frühbeck G, Burcelin R, Fernández-Real JM (2012). "Circulating lipopolysaccharide-binding protein (LBP) as a marker of obesity-related insulin resistance". Int J Obes (Lond). 36 (11): 1442–9. doi:10.1038/ijo.2011.256. PMID 22184060.
- Lepper PM, Schumann C, Triantafilou K, Rasche FM, Schuster T, Frank H, Schneider EM, Triantafilou M, von Eynatten M (2007). "Association of lipopolysaccharide-binding protein and coronary artery disease in men". J. Am. Coll. Cardiol. 50 (1): 25–31. doi:10.1016/j.jacc.2007.02.070. PMID 17601541.
- Ruiz AG, Casafont F, Crespo J, Cayón A, Mayorga M, Estebanez A, Fernadez-Escalante JC, Pons-Romero F (2007). "Lipopolysaccharide-binding protein plasma levels and liver TNF-alpha gene expression in obese patients: evidence for the potential role of endotoxin in the pathogenesis of non-alcoholic steatohepatitis". Obes Surg. 17 (10): 1374–80. doi:10.1007/s11695-007-9243-7. PMID 18000721.
- Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, Neyrinck AM, Fava F, Tuohy KM, Chabo C, Waget A, Delmée E, Cousin B, Sulpice T, Chamontin B, Ferrières J, Tanti JF, Gibson GR, Casteilla L, Delzenne NM, Alessi MC, Burcelin R (2007). "Metabolic endotoxemia initiates obesity and insulin resistance". Diabetes. 56 (7): 1761–72. doi:10.2337/db06-1491. PMID 17456850.
- Fei N, Zhao L (December 2012). "An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice". ISME J. 7 (4): 880–4. doi:10.1038/ismej.2012.153. PMC 3603399. PMID 23235292.
- Wicks, Ian P.; Howell, Meredith L.; Hancock, Tuesday; Kohsaka, Hitoshi; Olee, Tsaiwei; Carson, Dennis A. (March 1995). "Bacterial Lipopolysaccharide Copurifies with Plasmid DNA: Implications for Animal Models and Human Gene Therapy". Human Gene Therapy. 6 (3): 317–323. doi:10.1089/hum.1995.6.3-317. PMID 7779915.
- 16 December 2014. "The Detection of Endotoxins Via the LAL Test, the Chromogenic Method". Archived from the original on 29 March 2015. Retrieved 14 March 2015.
- Iwanaga S (2007). "Biochemical principle of Limulus test for detecting bacterial endotoxins". Proc. Jpn. Acad., Ser. B, Phys. Biol. Sci. 83 (4): 110–9. Bibcode:2007PJAB...83..110I. doi:10.2183/pjab.83.110. PMC 3756735. PMID 24019589.
- Ding JL, Ho B (2001). "A new era in pyrogen testing" (PDF). Trends Biotechnol. 19 (8): 277–81. doi:10.1016/s0167-7799(01)01694-8. PMID 11451451. Archived from the original (PDF) on 2 January 2014. Retrieved 2 January 2014.
External links
- Lipopolysaccharides at the US National Library of Medicine Medical Subject Headings (MeSH)