Plant litter
Litterfall, plant litter, leaf litter, tree litter, soil litter, or duff, is dead plant material (such as leaves, bark, needles, twigs, and cladodes) that have fallen to the ground. This detritus or dead organic material and its constituent nutrients are added to the top layer of soil, commonly known as the litter layer or O horizon ("O" for "organic"). Litter is an important factor in ecosystem dynamics, as it is indicative of ecological productivity and may be useful in predicting regional nutrient cycling and soil fertility.[1]

Characteristics and variability

Litterfall is characterized as fresh, undecomposed, and easily recognizable (by species and type) plant debris. This can be anything from leaves, cones, needles, twigs, bark, seeds/nuts, logs, or reproductive organs (e.g. the stamen of flowering plants). Items larger than 2 cm diameter are referred to as coarse litter, while anything smaller is referred to as fine litter or litter. The type of litterfall is most directly affected by ecosystem type. For example, leaf tissues account for about 70 percent of litterfall in forests, but woody litter tends to increase with forest age.[2] In grasslands, there is very little aboveground perennial tissue so the annual litterfall is very low and quite nearly equal to the net primary production.[3]
In soil science, soil litter is classified in three layers, which form on the surface of the O Horizon. These are the L, F, and H layers:[4]
- L – organic horizon characterized by relatively undecomposed plant material (described above).
- F – organic horizon found beneath L characterized by accumulation of partly decomposed organic matter.
- H – organic horizon below F characterized by accumulation of fully decomposed organic matter mostly indiscernible
The litter layer is quite variable in its thickness, decomposition rate and nutrient content and is affected in part by seasonality, plant species, climate, soil fertility, elevation, and latitude.[1] The most extreme variability of litterfall is seen as a function of seasonality; each individual species of plant has seasonal losses of certain parts of its body, which can be determined by the collection and classification of plant litterfall throughout the year, and in turn affects the thickness of the litter layer. In tropical environments, the largest amount of debris falls in the latter part of dry seasons and early during wet season.[5] As a result of this variability due to seasons, the decomposition rate for any given area will also be variable.
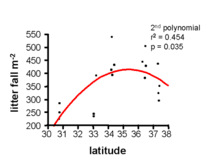
Latitude also has a strong effect on litterfall rates and thickness. Specifically, litterfall declines with increasing latitude. In tropical rainforests, there is a thin litter layer due to the rapid decomposition,[7] while in boreal forests, the rate of decomposition is slower and leads to the accumulation of a thick litter layer, also known as a mor.[3] Net primary production works inversely to this trend, suggesting that the accumulation of organic matter is mainly a result of decomposition rate.
Surface detritus facilitates the capture and infiltration of rainwater into lower soil layers. Soil litter protects soil aggregates from raindrop impact, preventing the release of clay and silt particles from plugging soil pores.[8] Releasing clay and silt particles reduces the capacity for soil to absorb water and increases cross surface flow, accelerating soil erosion. In addition soil litter reduces wind erosion by preventing soil from losing moisture and providing cover preventing soil transportation.
Organic matter accumulation also helps protect soils from wildfire damage. Soil litter can be completely removed depending on intensity and severity of wildfires and season.[9] Regions with high frequency wildfires have reduced vegetation density and reduced soil litter accumulation. Climate also influences the depth of plant litter. Typically humid tropical and sub-tropical climates have reduced organic matter layers and horizons due to year-round decomposition and high vegetation density and growth. In temperate and cold climates, litter tends to accumulate and decompose slower due to a shorter growing season.
Net primary productivity
Net primary production and litterfall are intimately connected. In every terrestrial ecosystem, the largest fraction of all net primary production is lost to herbivores and litterfall. Due to their interconnectedness, global patterns of litterfall are similar to global patterns of net primary productivity.[3]
Habitat and food
Litter provides habitat for a variety of organisms.
Plants

Certain plants are specially adapted for germinating and thriving in the litter layers. For example, bluebell (Hyacinthoides non-scripta) shoots puncture the layer to emerge in spring. Some plants with rhizomes, such as common wood sorrel (Oxalis acetosella) do well in this habitat.[7]
Detritivores and other decomposers


Many organisms that live on the forest floor are decomposers, such as fungi. Organisms whose diet consists of plant detritus, such as earthworms, are termed detritivores. The community of decomposers in the litter layer also includes bacteria, amoeba, nematodes, rotifer, tardigrades, springtails, cryptostigmata, potworms, insect larvae, mollusks, oribatid mites, woodlice, and millipedes.[7] Even some species of microcrustaceans, especially copepods (for instance Bryocyclops spp., Graeteriella spp.,Olmeccyclops hondo, Moraria spp.,Bryocamptus spp., Atheyella spp.)[10] live in moist leaf litter habitats and play an important role as predators and decomposers.[11]
The consumption of the litterfall by decomposers results in the breakdown of simple carbon compounds into carbon dioxide (CO2) and water (H2O), and releases inorganic ions (like nitrogen and phosphorus) into the soil where the surrounding plants can then reabsorb the nutrients that were shed as litterfall. In this way, litterfall becomes an important part of the nutrient cycle that sustains forest environments.
As litter decomposes, nutrients are released into the environment. The portion of the litter that is not readily decomposable is known as humus. Litter aids in soil moisture retention by cooling the ground surface and holding moisture in decaying organic matter. The flora and fauna working to decompose soil litter also aid in soil respiration. A litter layer of decomposing biomass provides a continuous energy source for macro- and micro-organisms.[12]
Larger animals
Numerous reptiles, amphibians, birds, and even some mammals rely on litter for shelter and forage. Amphibians such as salamanders and caecilians inhabit the damp microclimate underneath fallen leaves for part or all of their life cycle. This makes them difficult to observe. A BBC film crew captured footage of a female caecilian with young for the first time in a documentary that aired in 2008.[13] Some species of birds, such as the ovenbird of eastern North America for example, require leaf litter for both foraging and material for nests.[14] Sometimes litterfall even provides energy to much larger mammals, such as in boreal forests where lichen litterfall is one of the main constituents of wintering deer and elk diets.[15]
Nutrient cycle
During leaf senescence, a portion of the plant's nutrients are reabsorbed from the leaves. The nutrient concentrations in litterfall differ from the nutrient concentrations in the mature foliage by the reabsorption of constituents during leaf senescence.[3] Plants that grow in areas with low nutrient availability tend to produce litter with low nutrient concentrations, as a larger proportion of the available nutrients is reabsorbed. After senescence, the nutrient-enriched leaves become litterfall and settle on the soil below.
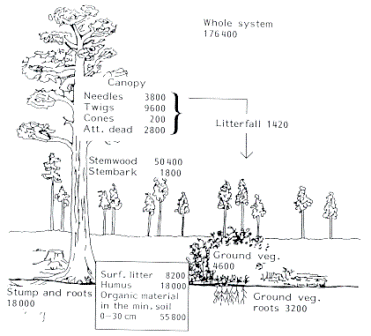
Litterfall is the dominant pathway for nutrient return to the soil, especially for nitrogen (N) and phosphorus (P). The accumulation of these nutrients in the top layer of soil is known as soil immobilization. Once the litterfall has settled, decomposition of the litter layer, accomplished through the leaching of nutrients by rainfall and throughfall and by the efforts of detritivores, releases the breakdown products into the soil below and therefore contributes to the cation exchange capacity of the soil. This holds especially true for highly weathered tropical soils.[17]
Leaching is the process by which cations such as iron (Fe) and aluminum (Al), as well as organic matter are removed from the litterfall and transported downward into the soil below. This process is known as podzolization and is particularly intense in boreal and cool temperate forests that are mainly constituted by coniferous pines whose litterfall is rich in phenolic compounds and fulvic acid.[3]
By the process of biological decomposition by microfauna, bacteria, and fungi, CO2 and H2O, nutrient elements, and a decomposition-resistant organic substance called humus are released. Humus composes the bulk of organic matter in the lower soil profile.[3]
The decline of nutrient ratios is also a function of decomposition of litterfall (i.e. as litterfall decomposes, more nutrients enter the soil below and the litter will have a lower nutrient ratio). Litterfall containing high nutrient concentrations will decompose more rapidly and asymptote as those nutrients decrease.[18] Knowing this, ecologists have been able to use nutrient concentrations as measured by remote sensing as an index of a potential rate of decomposition for any given area.[19] Globally, data from various forest ecosystems shows an inverse relationship in the decline in nutrient ratios to the apparent nutrition availability of the forest.[3]
Once nutrients have re-entered the soil, the plants can then reabsorb them through their roots. Therefore, nutrient reabsorption during senescence presents an opportunity for a plant's future net primary production use. A relationship between nutrient stores can also be defined as:
- annual storage of nutrients in plant tissues + replacement of losses from litterfall and leaching = the amount of uptake in an ecosystem
Collection and analysis
The main objectives of litterfall sampling and analysis are to quantify litterfall production and chemical composition over time in order to assess the variation in litterfall quantities, and hence its role in nutrient cycling across an environmental gradient of climate (moisture and temperature) and soil conditions.[20]
Ecologists employ a simple approach to the collection of litterfall, most of which centers around one piece of equipment, known as a litterbag. A litterbag is simply any type of container that can be set out in any given area for a specified amount of time to collect the plant litter that falls from the canopy above.

Litterbags are generally set in random locations within a given area and marked with GPS or local coordinates, and then monitored on a specific time interval. Once the samples have been collected, they are usually classified on type, size and species (if possible) and recorded on a spreadsheet.[22] When measuring bulk litterfall for an area, ecologists will weigh the dry contents of the litterbag. By this method litterfall flux can be defined as:
- litterfall (kg m−2 yr−1) = total litter mass (kg) / litterbag area (m2)[23]
The litterbag may also be used to study decomposition of the litter layer. By confining fresh litter in the mesh bags and placing them on the ground, an ecologist can monitor and collect the decay measurements of that litter.[7] An exponential decay pattern has been produced by this type of experiment: , where is the initial leaf litter and is a constant fraction of detrital mass.[3]



The mass-balance approach is also utilized in these experiments and suggests that the decomposition for a given amount of time should equal the input of litterfall for that same amount of time.
- litterfall = k(detrital mass)[3]
For study various groups from edaphic fauna you need a different mesh sizes in the litterbags[24]
Issues
Change due to invasive earthworms
In some regions of glaciated North America, earthworms have been introduced where they are not native. Non-native earthworms have led to environmental changes by accelerating the rate of decomposition of litter. These changes are being studied, but may have negative impacts on some inhabitants such as salamanders.[25]
Forest litter raking
Leaf litter accumulation depends on factors like wind, decomposition rate and species composition of the forest. The quantity, depth and humidity of leaf litter varies in different habitats. The leaf litter found in primary forests is more abundant, deeper and holds more humidity than in secondary forests. This condition also allows for a more stable leaf litter quantity throughout the year.[26] This thin, delicate layer of organic material can be easily affected by humans. For instance, forest litter raking as a replacement for straw in husbandry is an old non-timber practice in forest management that has been widespread in Europe since the seventeenth century.[27][28] In 1853, an estimated 50 Tg of dry litter per year was raked in European forests, when the practice reached its peak.[29] This human disturbance, if not combined with other degradation factors, could promote podzolisation; if managed properly (for example, by burying litter removed after its use in animal husbandry), even the repeated removal of forest biomass may not have negative effects on pedogenesis.[30]
See also
References
- Ochoa-Hueso, R; Delgado-Baquerizo, M; King, PTA; Benham, M; Arca, V; Power, SA (February 2019). "Ecosystem type and resource quality are more important than global change drivers in regulating early stages of litter decomposition". Soil Biology and Biochemistry. 129: 144–152. doi:10.1016/j.soilbio.2018.11.009.
- W. M. Lonsdale (1988). "Predicting the amount of litterfall in forests of the world". Annals of Botany. 61 (3): 319–324. doi:10.1093/oxfordjournals.aob.a087560.
- Schlesinger, William H. Biogeochemistry: An Analysis of Global Change. 2nd Edition. Academic Press. 108, 135, 152–158, 180–183, 191–194. (1997).
- "Soil Classification". Faculty of Land and Food Systems. The University of British Columbia. Retrieved March 20, 2012.
- A. V. Spain (1984). "Litterfall and the standing crop of litter in three tropical Australian rainforests". Journal of Ecology. 72 (3): 947–961. doi:10.2307/2259543. JSTOR 2259543.
- "Litter Fall in the North American Baldcypress Swamp Network, Illinois to Louisiana, 2003". Nwrc.usgs.gov. 2013-08-19. Retrieved 2014-04-09.
- Packham, J.R.; Harding, D.J.L.; Hilton, G.M.; Stuttard, R.A. (1992). Functional Ecology of Woodlands and Forests. London: Chapman & Hall. pp. 133–134, 246–247, 265. ISBN 0412439506.
- Chanasyk, D.S.; Whitson, I.R.; Mapfumo, E.; Burke, J.M.; Prepas, E.E. (2003). "The Impacts of Forest Harvest and Wildfire on Soils and Hydrology in Temperate Forests: A Baseline to Develop Hypotheses for the Boreal Plain". Journal of Environmental Engineering and Science. 2: S51–S62. doi:10.1139/S03-034.
- Ice, George G.; Neary, D.G.; Adams, P.W. (2004). "Effects of Wildfire on Soils and Watershed Processes" (PDF). Journal of Forestry. 102 (6): 16–20(5). Retrieved March 20, 2012.
- Fiers, Frank; Jocque, Merlijn (2013-03-20). "Leaf litter copepods from a cloud forest mountain top in Honduras (Copepoda: Cyclopidae, Canthocamptidae)". Zootaxa. 3630 (2): 270–290. doi:10.11646/zootaxa.3630.2.4. ISSN 1175-5334. PMID 26131511.
- Fiers, Frank Fiers; Ghenne, Véronique (January 2000). "Cryptozoic copepods from Belgium: Diversity and biogeographic implications". Belgian Journal of Zoology. 130 (1): 11–19.
- Bot, Alexandra (2005). The Importance of Soil Organic Matter. Rome: Food and Agriculture Organizations of the United Nations. pp. Chapter 3. ISBN 92-5-105366-9.
- Writer David Attenborough, Director Scott Alexander, Producer Hilary Jeffkins (2008-02-11). "Land Invaders". Life in Cold Blood. BBC. BBC One.
- Dunn, Jon; Garrett, Kimball (1997). Warblers. New York: Peterson Field Guides. p. 451. ISBN 0-395-78321-6.
- Richard L. Ward & C. Les Marcum (2005). "Lichen litterfall consumption by wintering deer and elk in western Montana". Journal of Wildlife Management. 69 (3): 1081–1089. doi:10.2193/0022-541X(2005)069[1081:LLCBWD]2.0.CO;2. JSTOR 3803347.
- Breymeyer, A.I., B. Berg, S.T. Gower, & D. Johnson. “Temperate Coniferous Forests” Scientific Committee on Problems of the Environment (SCOPE). Vol. 56: Global Change: Effects on Coniferous Forests and Grasslands Carbon Budget, Ch. 3. (1996).
- J. Chave, D. Navarrete, S. Almeida, E. Álvarez, L. E. O. C. Aragão, D. Bonal, P. Châtelet, J. E. Silva-Espejo, J.-Y. Goret, P. von Hildebrand, E. Jiménez, S. Patiño, M. C. Peñuela, O. L. Phillips, P. Stevenson & Y. Malhi (2009). "Regional and seasonal patterns of litterfall in tropical South America" (PDF). Biogeosciences. 7 (1): 43–55. doi:10.5194/bg-7-43-2010.CS1 maint: multiple names: authors list (link)
- Scott D. Bridgham; John Pastor; Charles A. McClaugherty & Curtis J. Richardson (1995). "Nutrient-use efficiency: a litterfall index, a model, and a test along a nutrient-availability gradient in North Carolina peatlands" (PDF). The American Naturalist. 145 (1): 1–21. doi:10.1086/285725. S2CID 84467103. Archived from the original (PDF) on 2011-08-13.
- Melillo, J.M., & J.R. Gosz. “Interactions of Biogeochemical Cycles in Forest Ecosystems” Scientific Committee on Problems of the Environment (SCOPE). Vol. 21: The Major Biogeochemical Cycles and Their Interactions, Ch. 6. (1983).
- Simmons, Jeffrey A. “Measuring Litterfall Flux.” West Virginia Wesleyan College (2003).
- "Spatial variations of nitrogen deposition and its effect on forest biochemical processes". Forest Research. Retrieved March 27, 2011.
- Estrella, Stephanie. “Standard Operating Procedures for Litterfall Collection, Processing, and Analysis: Version 2.0.” Washington State Department of Ecology. (2008).
- Bastrup-Birk, A., & Nathalie Bréda. “Report on Sampling and Analysis of Litterfall” United Nations Economic Commission for Europe Convention on Long-Range Transboundary Air Pollution: International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests. (2004).
- Castro-Huerta, R., Falco, L., Sandler, R., Coviella, C. (2015). "Differential contribution of soil biota groups to plant litter decomposition as mediated by soil use". PeerJ. 3: e826. doi:10.7717/peerj.826. PMC 4359044. PMID 25780777.CS1 maint: multiple names: authors list (link)
- Maerz, John C.; Nuzzo, Victoria A.; Blossey, Bernd (2009). "Declines in Woodland Salamander Abundance Associated with Non-Native Earthworm and Plant Invasions" (PDF). Conservation Biology. 23 (4): 975–981. doi:10.1111/j.1523-1739.2009.01167.x. PMID 19236449. Retrieved 28 April 2012.
- Barrientos, Zaidett (2012). "Dynamics of leaf litter humidity, depth and quantity: two restoration strategies failed to mimic ground microhabitat conditions of a low montane and premontane forest in Costa Rica" (PDF). Revista de Biología Tropical. 60 (3): 1041–1053. doi:10.15517/rbt.v60i3.1756. PMID 23025078.
- Bürgi, M., Gimmi, U. (2007). "Three objectives of historical ecology: the case of litter collecting in Central European forests". Landscape Ecology. 22: 77–87. doi:10.1007/s10980-007-9128-0. hdl:20.500.11850/58945. S2CID 21130814.CS1 maint: multiple names: authors list (link)
- Gimmi, U., Poulter, B., Wolf, A., Portner, H., Weber, P., Bürgi, M. (2013). "Soil carbon pools in Swiss forests show legacy effects from historic forest litter raking" (PDF). Landscape Ecology. 28 (5): 385–846. doi:10.1007/s10980-012-9778-4. hdl:20.500.11850/66782. S2CID 16930894.CS1 maint: multiple names: authors list (link)
- McGrath, M.J.; et al. (2015). "Reconstructing European forest management from 1600 to 2010". Biogeosciences. 12 (14): 4291–4316. Bibcode:2015BGeo...12.4291M. doi:10.5194/bg-12-4291-2015.
- Scalenghe, R, Minoja, A.P., Zimmermann, S., Bertini, S. (2016). "Consequence of litter removal on pedogenesis: A case study in Bachs and Irchel (Switzerland)". Geoderma. 271: 191–201. Bibcode:2016Geode.271..191S. doi:10.1016/j.geoderma.2016.02.024.CS1 maint: multiple names: authors list (link)
