Sucrase-isomaltase
Sucrase-isomaltase (SI) is a glucosidase enzyme located on the brush border of the small intestine. It is a dual-function enzyme with two GH31 domains, one serving as the isomaltase, the other as a sucrose alpha-glucosidase.[5][6][7] It has preferential expression in the apical membranes of enterocytes.[8] The enzyme’s purpose is to digest dietary carbohydrates such as starch, sucrose and isomaltose. By further processing the broken-down products, energy in the form of ATP can be generated.[9]


Structure
Sucrase-isomaltase consists of two enzymatic subunits: sucrase and isomaltase. The subunits originate from a polypeptide precursor, pro-SI. By heterodimerizing the two subunits, the sucrase-isomaltase complex is formed.[11] The enzyme is anchored in the intestinal brush border membrane by a hydrophobic segment located near the N-terminal of the isomaltase subunit.[12] Before the enzyme is anchored to the membrane, pro-SI is mannose-rich and glycosylated; it moves from the ER to the Golgi, where it becomes a protein complex that is N- and O- glycosylated. The O-linked glycosylation is necessary to target the protein to the apical membrane.[13][14] In addition, there is a segment that is both O-linked glycosylated and Ser/Thr-rich.[15] A similarly-arranged enzyme is the maltase-glucoamylase, also a member of GH31.
Sucrase-isomaltase is composed of duplicated catalytic domains, N- and C-terminal. Each domain displays overlapping specificities. Scientists have discovered the crystal structure for N-terminal human sucrase-isomaltase (ntSI) in apo form to 3.2 Å and in complex with the inhibitor kotalanol to 2.15 Å resolution.[10] Sucrase-isomaltase’s mechanism results in a net retention of configuration at the anomeric center.[10]
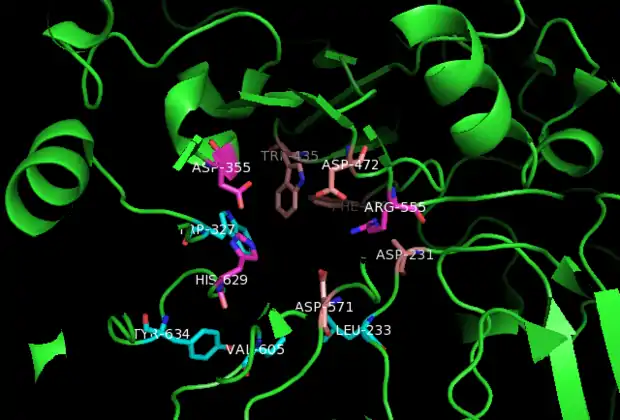
The crystal structure shows that sucrase-isomaltase exists as a monomer. The researchers claim that the observance of SI dimers is dependent on experimental conditions.[10] ntSI’s four monomers, A, B, C, and D are included in the crystal asymmetric unit and have identical active sites. The active site is composed of a shallow-substrate binding pocket including -1 and +1 subsites. The non-reducing end of substrates binds to the pocket. While the non-reducing sugar ring has interactions with the buried -1 subsite, the reducing ring has interactions with the surface exposed +1 subsite.[10]
The interactions between the active site of sucrase-isomaltase and the following compounds have been identified:
- Man2GlcNAc2 glycan: Within the active site, Man2GlcNAc2 hydrogen bonds with hydroxyl side chains of Asp231 and Asp571. Furthermore, hydrophobic interactions with Leu233, Trp327, Trp435, Phe479, Val605, and Tyr634 provide additional stabilization for Man2GlcNAc2.[10]
- Kotalonal, the inhibitor: It interacts with the catalytic nucleophile Asp472 and acid base catalyst Asp571. In addition, ntSI residues His629, Asp355, Arg555, Asp231, Trp435, and Phe479 bind to the substrate.[10]
Currently, there are no crystal structures of ntSI in complex with an α-1,6-linked substrate or inhibitor analogue. In order to predict isomaltose binding in sucrase-isomaltase structure, a model was produced by hand. Within the -1 subsite, isomaltose’s non-reducing glucose ring was aligned to that of acarbose.[10]
Not only has the structure of human sucrase-isomaltase been studied, but also sucrase-isomaltase’s structure in sea lions and pigs have also been analyzed.[6][16][17]
Disease relevance
A deficiency is responsible for sucrose intolerance. Congenital sucrase-isomaltase deficiency (CSID), also called genetic sucrase-isomaltase deficiency (GSID), and sucrose intolerance, is a genetic, intestinal disorder that is caused by a reduction or absence of sucrase and isomaltase[14] Explanations for GSID include:
- Mutations C1229Y and F1745C, which are present in the sucrase domain of SI, block SI path to anchor in the cell’s aprical membrane but does not impact protein folding or isomaltase activity.[14]
- Substitution of a cysteine by an arginine at amino acid residue 635 in the isomaltase subunit of SI was present in the cDNA encoding for a patient with CSID. SIC635R had an altered folding pattern, which influenced the sorting profile and increased the turnover rate.[18]
- A factor that can be attributed to congenital sucrase-isomaltase deficiency is the retention of SI in the cis-Golgi. This inability to transport is a result of a glutamine to proline substitution at amino acid residue 1098 of the sucrase subunit.[19][20]
Furthermore, a relationship between mutations in sucrase-isomaltase and chronic lymphocytic leukemia (CLL) has been identified. These mutations cause a loss of enzyme function by blocking the biosynthesis of SI at the cell surface.[8]
See also
References
- GRCh38: Ensembl release 89: ENSG00000090402 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000027790 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Hauri HP, Quaroni A, Isselbacher KJ (October 1979). "Biogenesis of intestinal plasma membrane: posttranslational route and cleavage of sucrase-isomaltase". Proceedings of the National Academy of Sciences of the United States of America. 76 (10): 5183–6. doi:10.1073/pnas.76.10.5183. PMC 413104. PMID 291933.
- Sjöström H, Norén O, Christiansen L, Wacker H, Semenza G (December 1980). "A fully active, two-active-site, single-chain sucrase.isomaltase from pig small intestine. Implications for the biosynthesis of a mammalian integral stalked membrane protein". The Journal of Biological Chemistry. 255 (23): 11332–8. PMID 7002920.
- Rodriguez IR, Taravel FR, Whelan WJ (September 1984). "Characterization and function of pig intestinal sucrase-isomaltase and its separate subunits". European Journal of Biochemistry / FEBS. 143 (3): 575–82. doi:10.1111/j.1432-1033.1984.tb08408.x. PMID 6479163.
- Rodríguez D, Ramsay AJ, Quesada V, Garabaya C, Campo E, Freije JM, López-Otín C (June 2013). "Functional analysis of sucrase-isomaltase mutations from chronic lymphocytic leukemia patients". Human Molecular Genetics. 22 (11): 2273–82. doi:10.1093/hmg/ddt078. PMID 23418305.
- Berg, J. M. et al. Biochemistry, 7th Ed. W.H. Freeman and Company: New York, 2012.
- Sim L, Willemsma C, Mohan S, Naim HY, Pinto BM, Rose DR (June 2010). "Structural basis for substrate selectivity in human maltase-glucoamylase and sucrase-isomaltase N-terminal domains". The Journal of Biological Chemistry. 285 (23): 17763–70. doi:10.1074/jbc.M109.078980. PMC 2878540. PMID 20356844.
- "SI sucrase-isomaltase (alpha-glucosidase) [Homo sapiens (human)] - Gene - NCBI".
- Brunner, J.; Hauser, H.; Braun, H.; Wilson, K. J.; Wacker, H.; O'Neill, B.; Semenza, G. (1979). "The mode of association of the enzyme complex sucrase-isomaltase with the intestinal brush border membrane" (PDF). J. Biol. Chem. 254 (6): 1821–8. PMID 422555.
- Naim HY, Sterchi EE, Lentze MJ (May 1988). "Biosynthesis of the human sucrase-isomaltase complex. Differential O-glycosylation of the sucrase subunit correlates with its position within the enzyme complex". The Journal of Biological Chemistry. 263 (15): 7242–53. PMID 3366777.
- Alfalah M, Keiser M, Leeb T, Zimmer KP, Naim HY (March 2009). "Compound heterozygous mutations affect protein folding and function in patients with congenital sucrase-isomaltase deficiency". Gastroenterology. 136 (3): 883–92. doi:10.1053/j.gastro.2008.11.038. PMID 19121318.
- Hunziker W, Spiess M, Semenza G, Lodish HF (July 1986). "The sucrase-isomaltase complex: primary structure, membrane-orientation, and evolution of a stalked, intrinsic brush border protein". Cell. 46 (2): 227–34. doi:10.1016/0092-8674(86)90739-7. PMID 3755079. S2CID 8207969.
- Wacker H, Aggeler R, Kretchmer N, O'Neill B, Takesue Y, Semenza G (April 1984). "A two-active site one-polypeptide enzyme: the isomaltase from sea lion small intestinal brush-border membrane. Its possible phylogenetic relationship with sucrase-isomaltase". The Journal of Biological Chemistry. 259 (8): 4878–84. PMID 6715326.
- Galand G (1989). "Brush border membrane sucrase-isomaltase, maltase-glucoamylase and trehalase in mammals. Comparative development, effects of glucocorticoids, molecular mechanisms, and phylogenetic implications". Comparative Biochemistry and Physiology. B, Comparative Biochemistry. 94 (1): 1–11. doi:10.1016/0305-0491(89)90002-3. PMID 2513162.
- Keiser M, Alfalah M, Pröpsting MJ, Castelletti D, Naim HY (May 2006). "Altered folding, turnover, and polarized sorting act in concert to define a novel pathomechanism of congenital sucrase-isomaltase deficiency". The Journal of Biological Chemistry. 281 (20): 14393–9. doi:10.1074/jbc.M513631200. PMID 16543230.
- Pröpsting MJ, Kanapin H, Jacob R, Naim HY (June 2005). "A phenylalanine-based folding determinant in intestinal sucrase-isomaltase that functions in the context of a quality control mechanism beyond the endoplasmic reticulum". Journal of Cell Science. 118 (Pt 12): 2775–84. doi:10.1242/jcs.02364. PMID 15944403.
- Pröpsting MJ, Jacob R, Naim HY (May 2003). "A glutamine to proline exchange at amino acid residue 1098 in sucrase causes a temperature-sensitive arrest of sucrase-isomaltase in the endoplasmic reticulum and cis-Golgi". The Journal of Biological Chemistry. 278 (18): 16310–4. doi:10.1074/jbc.C300093200. PMID 12624106.
External links
- Structure and evolution of the mammalian maltase-glucoamylase and sucrase-isomaltase
- Sucrase-isomaltase+complex at the US National Library of Medicine Medical Subject Headings (MeSH)