Anomalocaris
Anomalocaris ("unlike other shrimp", or "abnormal shrimp") is an extinct genus of radiodont (anomalocaridid), an order of animals thought to be closely related to ancestral arthropods. The first fossils of Anomalocaris were discovered in the Ogygopsis Shale by Joseph Frederick Whiteaves, with more examples found by Charles Doolittle Walcott in the Burgess Shale.[2] Originally several fossilized parts discovered separately (the mouth, frontal appendages and trunk) were thought to be three separate creatures, a misapprehension corrected by Harry B. Whittington and Derek Briggs in a 1985 journal article.[2][3] Anomalocaris is thought to be one of the earliest examples of an apex predator, though others have been found in older Cambrian lagerstätten deposits.
| Anomalocaris | |
|---|---|
 | |
| Image of the first complete Anomalocaris fossil found, residing in the Royal Ontario Museum | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | †Dinocaridida |
| Order: | †Radiodonta |
| Family: | †Anomalocarididae |
| Genus: | †Anomalocaris Whiteaves, 1892 |
| Species | |
| |
| Synonyms | |
|
Defunct species:[1]
| |
Discovery
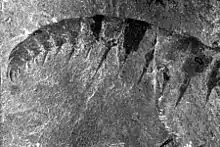
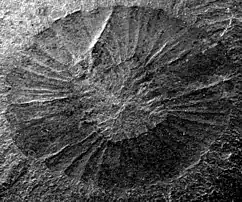
Anomalocaris has been misidentified several times, in part due to its makeup of a mixture of mineralized and unmineralized body parts; the mouth and frontal appendage was considerably harder and more easily fossilized than the delicate body.[4] Anomalocaris fossils were first collected in 1886[5] or 1888[6] by Richard G. McConnell of the Geological Survey of Canada. The specimens were described and named in 1892 by GSC paleontologist Joseph Frederick Whiteaves.[6] The specimens are now known to represent isolated frontal appendages, but Whiteaves interpreted them as the abdomens of phyllocarid crustaceans. Noting its unusual anatomy for the abdomen of a crustacean, Whiteaves gave it the name Anomalocaris, meaning "unlike other shrimps". In 1928, Danish paleontologist Kai Henriksen proposed that Tuzoia, a Burgess Shale arthropod then known only from the carapace, represented the missing front half of Anomalocaris.[5] The artists Elie Cheverlange and Charles R. Knight followed this interpretation in their depictions of Anomalocaris.[5]
Unknown to scientists at the time, the body parts of relatives of Anomalocaris had already been described but not recognized as such. The first fossilized anomalocaridid mouth was discovered by Charles Doolittle Walcott, who mistook it for a jellyfish and placed it in the genus Peytoia. Walcott also discovered a frontal appendage but failed to realize the similarities to Whiteaves' discovery and instead identified it as feeding appendage or tail of the extinct Sidneyia.[4] In the same publication in which he named Peytoia, Walcott named Laggania, a taxon that he interpreted as a holothurian.
In 1966, the Geological Survey of Canada began a comprehensive revision of the Burgess Shale fossil record, led by Cambridge University paleontologist Harry B. Whittington.[5] In the process of this revision, Whittington and his students Simon Conway Morris and Derek Briggs would discover the true nature of Anomalocaris and its relatives, but not without contributing to the history of misinterpretations first.[4] In 1978, Conway Morris recognized that the mouthparts of Laggania were identical to Peytoia, but concluded that Laggania was a composite fossil made up of Peytoia and the sponge Corralio undulata.[7] In 1979, Briggs recognized that the fossils of Anomalocaris were appendages, not abdomens, and proposed that they were the walking legs of a giant arthropod, and that the feeding appendage Walcott had assigned to Sidneyia was the feeding appendage of Anomalocaris.[8] Later, while clearing what he thought was an unrelated specimen, Harry B. Whittington removed a layer of covering stone to discover the unequivocally connected frontal appendage thought to be a shrimp tail and mouth thought to be a jellyfish.[4][2] Whittington linked the two species, but it took several more years for researchers to realize that the continuously juxtaposed Peytoia, Laggania and frontal appendage actually represented a single, enormous creature. The two genera are now placed into the order Radiodonta, which commonly known as radiodont or anomalocaridid. Because Peytoia was named first, it became the correct name for the entire animal. The original frontal appendage, however, came from a larger species distinct from Peytoia and "Laggania", which retains the name Anomalocaris.[9]
In 2011, six fossils of compound eyes, the first for Anomalocaris, were recovered from a paleontological dig at Emu Bay on Kangaroo Island, Australia, proving that Anomalocaris was indeed an arthropod as had been suspected. The find also indicated that advanced arthropod eyes had evolved very early, before the evolution of jointed legs or hardened exoskeletons.[10]
Stephen Jay Gould cites Anomalocaris as one of the fossilized extinct species he believed to be evidence of a much more diverse set of phyla that existed in the Cambrian Period,[4] discussed in his book Wonderful Life, a conclusion disputed by other paleontologists.[2]
Anatomy
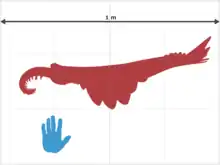
For the time in which it lived, Anomalocaris was gigantic, up to 1 metre (3.3 feet) long.[2] It propelled itself through the water by undulating the flexible flaps on the sides of its body.[11] Each flap sloped below the one more posterior to it,[12] and this overlapping allowed the lobes on each side of the body to act as a single "fin", maximizing the swimming efficiency.[11] The construction of a remote-controlled model showed this mode of swimming to be intrinsically stable,[13] implying that Anomalocaris would not have needed a complex brain to manage balance while swimming. The body was widest between the third and fifth lobe and narrowed towards the tail; it was thought to have at least 11 pairs of flaps in total. It is difficult to distinguish lobes near the tail, making an accurate count difficult.[12] Later studies reveal that it had 13 pairs on the trunk region and additional 3 pairs of small flaps on the neck region.[14]

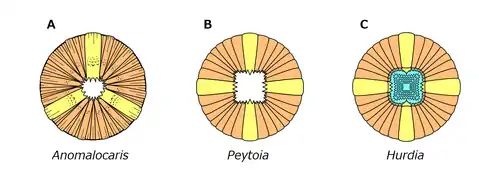
Anomalocaris had an unusual disk-like mouth known as oral cone. The oral cone was composed of several plates organized triradially. Three of the plates were quite large. Three to four medium sized plates could be found between each of the large plates, and several small, wrinkled plates between them.[9] Such oral cone is very different to those of a typical hurdiid radiodont like Peytoia and Hurdia, which is smooth and tetraradial.[9] The mouth resembled a pineapple ring with the center replaced by a series of serrated prongs.[2] The mouth could constrict to crush prey, but never completely close; there was always an opening about 5 millimeters in diameter when the mouth was shut; a centimeter in exceptionally large individuals. Two large frontal appendage (up to 18 centimetres (7.1 inches) in length when extended)[4] were positioned in front of the mouth, at the front of the head. As a shared character across radiodonts,[15] Anomalocaris also possessed three sclerites on the top and side of its head. The top one known as a head shield, dorsal carapace or H-element. it was shaped like an oval, with a distinct rim on the outer edge.[14] The remaining two lateral sclerites known as P-elements also ovoid, but connected by a bar-like outgrowth.[15] The P-elements were previously misinterpreted as two huge compound eyes.[14][15]
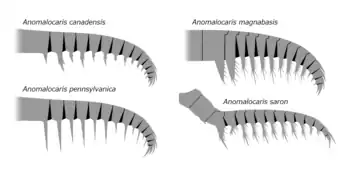
Anomalocaris canadensis possessed 14 segments (podomeres) on each frontal appendage, almost each one tipped with two barb-like ventral spines (endites). The ventral spines themselves were both equipped with 2 smaller auxiliary spines, which branches off from the main spine at the center of its length.[3] The tail was large and fan-shaped, composed of three pairs of large, lateral fin-shaped lobes and one terminal lobe-like tailpiece.[14] Along with undulations of the lobes, it was probably used to propel the creature through Cambrian waters.[2][11][16] The gills of the animal, in the form of long, thin, hair-like structures known as lanceolate blades, arranged in a row which forming setal blades. The setal blades were attached by their margin to the top side of the animal, two setal blades per body segment. A divide ran down the middle, separating the gills.[14]
Based on fossilized eyes from the Emu Bay Shale, which belonged to Anomalocaris briggsi (which, based on morphological and phylogenetic evidence, was more closely related to Tamisiocaris), the eyes of Anomalocaris were 30 times more powerful than those of trilobites, long thought to have had the most advanced eyes of any contemporary species. With 16,000 lenses, the resolution of the 3-centimetre-wide (1.2 in) eyes would have been rivalled only by that of the modern dragonfly, which has 28,000 lenses in each eye.[10] Additionally, Anomalocaris had dichromatic colour vision.[17]
Paleobiology
Diet

A long-standing view holds that Anomalocaris fed on hard-bodied animals, including trilobites—making it one of the first predators—and its mid-gut glands strongly suggest a predatory lifestyle.[18] However, its ability to penetrate mineralised shells has been questioned.[19] Some Cambrian trilobites have been found with round or W-shaped "bite" marks, which were identified as being the same shape as the mouthparts of Anomalocaris.[20] Stronger evidence that Anomalocaris ate trilobites comes from coprolite, which contain trilobite parts and are so large that the anomalocarids are the only known organism from that period large enough to have produced them.[20] However, since Anomalocaris lacks any mineralised tissue, it seemed unlikely that it would be able to penetrate the hard, calcified shell of trilobites.[20] Rather, the coprolites may have been produced by a different organisms, such as the trilobites of the genus Redlichia.[21]
The lack of wear on anomalocaridid mouthparts suggests they did not come into regular contact with mineralised trilobite shells, and were possibly better suited to feeding on smaller, soft-bodied organisms by suction, since they would have experienced structural failure if they were used against the armour of trilobites.[19][21] A. canadensis may have been capable of feeding on organisms with hard exoskeletons due to the short, robust spines on its appendages.[21] However, as opposed to Peytoia whose mouthpiece is more rectangular with short protruding spines, the mouthpiece of A. canadensis has a smaller and more irregular diamond-shaped opening, not permitting strong biting motions, and indicating a suction-feeding behavior to suck in softer organisms.[9]

Phylogenetic analyses have also found A. briggsi to belong to the closely related, filter-feeding family Cetiocaridae, which suggests that may have been a filter-feeder as well.[22][23]
One suggested possibility is that anomalocaridids fed by grabbing one end of their prey in their jaws while using their appendages to quickly rock the other end of the animal back and forth. This produced stresses that exploited the weaknesses of arthropod cuticles, causing the prey's exoskeleton to rupture and allowing the predator to access its innards.[20] This behaviour was originally thought to have provided an evolutionary pressure for trilobites to roll up, to avoid being flexed until they snapped.[20]
Anomalocaris magnabasis, a large species of Anomalocaris discovered south of the Burgess Shale, had spinules projecting from the spine base. These small, thin spines would have allowed Anomalocaris magnabasis to be a more effective hunter of soft bodied prey, able to pick up and quickly ensnare priapulids and other animals that lacked a hard shell.[24]
Paleoecology
Anomalocaris canadensis lived in the Burgess Shale in relatively great numbers, though comparable fossils have been found elsewhere, suggesting a more expansive range over the Laurentian continent.[25] In the Burgess Shale, Anomalocaris is more common in the older sections, notably the Mount Stephen trilobite beds. However, in the younger sections, such as the Phyllopod bed, Anomalocaris could reach much greater sizes; roughly twice the size of its older, trilobite bed relatives. These rare giant specimens have previously been referred to a separate species, Anomalocaris gigantea; however, the validity of this species has been called into question, and is currently dubious.[26]
Other species attributed to Anomalocaris live in vastly different environments. Anomalocaris saron and Anomalocaris kunmingensis lived in the Maotianshan Shales, a shallow tropical sea in what is now modern China. Anomalocaris briggsi lived in a comparable environment; the shallow, tropical waters of Cambrian Australia. The Maotianshan Shale and the Emu Bay Shale are very close in proximity, being separated by a small landmass, far from the Burgess Shale.[27]
See also
- 8564 Anomalocaris, an asteroid named after this animal
- Cambrian explosion
- Opabinia
- Wiwaxia
- Aegirocassis, a giant filter-feeding radiodont from Ordovician Morocco
Footnotes
- Daley AC, Peel JS (2010). "A Possible Anomalocaridid from the Cambrian Sirius Passet Lagerstätte, North Greenland". Journal of Paleontology. 84 (2): 352–355. doi:10.1666/09-136R1.1.
- Conway Morris S (1998). The crucible of creation: the Burgess Shale and the rise of animals. Oxford [Oxfordshire]: Oxford University Press. pp. 56–9. ISBN 978-0-19-850256-2.
- Whittington HB, Briggs DE (1985). "The largest Cambrian animal, Anomalocaris, Burgess Shale, British Columbia". Philosophical Transactions of the Royal Society B. 309 (1141): 569–609. Bibcode:1985RSPTB.309..569W. doi:10.1098/rstb.1985.0096.
- Gould SJ (1989). Wonderful life: the Burgess Shale and the nature of history. New York: W.W. Norton. pp. 194–206. ISBN 978-0-393-02705-1.
- Collins, Desmond (1996). "The "Evolution" of Anomalocaris and Its Classification in the Arthropod Class Dinocarida (nov.) and Order Radiodonta (nov.)". Journal of Paleontology. 70 (2): 280–293. JSTOR 1306391.
- Whiteaves, J. F. (1892). "Description of a new genus and species of phyllocarid Crustacea from the Middle Cambrian of Mount Stephen, B. C.". The Canadian Record of Science. 5 (4).
- Conway Morris, S. (1978). "Laggania cambria Walcott: A Composite Fossil". Journal of Paleontology. 52 (1): 126–131. JSTOR 1303799.
- Briggs, D. E. G. (1979). "Anomalocaris, the largest known Cambrian arthropod". Palaeontology. 22 (3): 631–664.
- Daley AC, Bergström J (June 2012). "The oral cone of Anomalocaris is not a classic peytoia". Die Naturwissenschaften. 99 (6): 501–4. Bibcode:2012NW.....99..501D. doi:10.1007/s00114-012-0910-8. PMID 22476406.
- Paterson JR, García-Bellido DC, Lee MS, Brock GA, Jago JB, Edgecombe GD (December 2011). "Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes". Nature. 480 (7376): 237–40. Bibcode:2011Natur.480..237P. doi:10.1038/nature10689. PMID 22158247.
- Usami Y (January 2006). "Theoretical study on the body form and swimming pattern of Anomalocaris based on hydrodynamic simulation". Journal of Theoretical Biology. 238 (1): 11–7. doi:10.1016/j.jtbi.2005.05.008. PMID 16002096.
- Whittington HB, Briggs DE (1985). "The Largest Cambrian Animal, Anomalocaris, Burgess Shale, British Columbia". Philosophical Transactions of the Royal Society B (free full text). 309 (1141): 569–609. Bibcode:1985RSPTB.309..569W. doi:10.1098/rstb.1985.0096.
- Briggs DE (May 1994). "Giant predators from the cambrian of china". Science. 264 (5163): 1283–4. Bibcode:1994Sci...264.1283B. doi:10.1126/science.264.5163.1283. PMID 17780843.
- Daley AC, Edgecombe GD (January 2014). "Morphology of Anomalocaris canadensis from the Burgess Shale". Journal of Paleontology. 88 (01): 68–91. doi:10.1666/13-067.
- Moysiuk J, Caron JB (August 2019). "A new hurdiid radiodont from the Burgess Shale evinces the exploitation of Cambrian infaunal food sources". Proceedings. Biological Sciences. 286 (1908): 20191079. doi:10.1098/rspb.2019.1079. PMC 6710600. PMID 31362637.
- "The Anomalocaris homepage". Archived from the original on 4 April 2008. Retrieved 2008-03-20.
- Fleming JF, Kristensen RM, Sørensen MV, Park TS, Arakawa K, Blaxter M, et al. (December 2018). "Molecular palaeontology illuminates the evolution of ecdysozoan vision". Proceedings. Biological Sciences. 285 (1892): 20182180. doi:10.1098/rspb.2018.2180. PMC 6283943. PMID 30518575.
- Vannier J, Liu J, Lerosey-Aubril R, Vinther J, Daley AC (May 2014). "Sophisticated digestive systems in early arthropods". Nature Communications. 5 (1): 3641. Bibcode:2014NatCo...5E3641V. doi:10.1038/ncomms4641. PMID 24785191.
- Hagadorn JW (August 2009). "Taking a Bite out of Anomalocaris" (PDF). In Smith MR, O'Brien LJ, Caron J (eds.). Abstract Volume. International Conference on the Cambrian Explosion (Walcott 2009). Toronto, Ontario, Canada: The Burgess Shale Consortium (published 31 July 2009). ISBN 978-0-9812885-1-2.
- Nedin C (1999). "Anomalocaris predation on nonmineralized and mineralized trilobites". Geology. 27 (11): 987–990. Bibcode:1999Geo....27..987N. doi:10.1130/0091-7613(1999)027<0987:APONAM>2.3.CO;2.
- Daley AC, Paterson JR, Edgecombe GD, García-Bellido DC, Jago JB (2013). Donoghue P (ed.). "New anatomical information on Anomalocaris from the Cambrian Emu Bay Shale of South Australia and a reassessment of its inferred predatory habits". Palaeontology: n/a. doi:10.1111/pala.12029.
- Vinther J, Stein M, Longrich NR, Harper DA (March 2014). "A suspension-feeding anomalocarid from the Early Cambrian" (PDF). Nature. 507 (7493): 496–9. Bibcode:2014Natur.507..496V. doi:10.1038/nature13010. PMID 24670770.
- Van Roy P, Daley AC, Briggs DE (June 2015). "Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps". Nature. 522 (7554): 77–80. Bibcode:2015Natur.522...77V. doi:10.1038/nature14256. PMID 25762145.
- Pates S, Daley AC, Edgecombe GD, Cong P, Lieberman BS (2019). "Systematics, preservation and biogeography of radiodonts from the southern Great Basin, USA, during the upper Dyeran (Cambrian Series 2, Stage 4)". Papers in Palaeontology. 0. doi:10.1002/spp2.1277.
- Lerosey-Aubril R, Hegna TA, Babcock LE, Bonino E, Kier C (2014-05-19). "Arthropod appendages from the Weeks Formation Konservat-Lagerstätte: new occurrences of anomalocaridids in the Cambrian of Utah, USA". Bulletin of Geosciences: 269–282. doi:10.3140/bull.geosci.1442.
- "Anomalocaris, the largest known Cambrian arthropod | The Palaeontological Association". www.palass.org. Retrieved 2019-09-16.
- Jeanes J. "Mapping the world's Burgess Shale-type deposits". www.virtualmuseum.ca. Retrieved 2019-09-16.
External links
| Wikimedia Commons has media related to Anomalocaris. |
- "Anomalocaris canadensis". Burgess Shale Fossil Gallery. Virtual Museum of Canada. 2011.
- Anomalocaris 'homepage' with swimming animation
- Burgess Shale: Anomalocaris canadensis (proto-arthropod), Smithsonian.
