Jurassic
The Jurassic (/dʒʊˈræs.sɪk/ juu-RASS-ik[1]) is a geologic period and system that spanned 56 million years from the end of the Triassic Period 201.3 million years ago (Mya) to the beginning of the Cretaceous Period 145 Mya.[note 1] The Jurassic constitutes the middle period of the Mesozoic Era. The Jurassic is named after the Jura Mountains in the European Alps, where limestone strata from the period were first identified.
| Jurassic | |
|---|---|
| 201.3 ± 0.2 – ~145.0 Ma | |
| Chronology | |
Key events in the Jurassic -200 — – -195 — – -190 — – -185 — – -180 — – -175 — – -170 — – -165 — – -160 — – -155 — – -150 — – -145 — – how to approximate the timescale of key Jurassic events. Vertical axis: millions of years ago. | |
| Etymology | |
| Name formality | Formal |
| Usage information | |
| Celestial body | Earth |
| Regional usage | Global (ICS) |
| Time scale(s) used | ICS Time Scale |
| Definition | |
| Chronological unit | Period |
| Stratigraphic unit | System |
| Time span formality | Formal |
| Lower boundary definition | First appearance of the Ammonite Psiloceras spelae tirolicum. |
| Lower boundary GSSP | Kuhjoch section, Karwendel mountains, Northern Calcareous Alps, Austria 47.4839°N 11.5306°E |
| GSSP ratified | 2010 |
| Upper boundary definition | Not formally defined |
| Upper boundary definition candidates |
|
| Upper boundary GSSP candidate section(s) | None |
| Atmospheric and climatic data | |
| Mean atmospheric O 2 content | c. 26 vol % (130 % of modern) |
| Mean atmospheric CO 2 content | c. 1950 ppm (7 times pre-industrial) |
| Mean surface temperature | c. 16.5 °C (3 °C above modern) |
The start of the period was marked by the major Triassic–Jurassic extinction event. Two other extinction events occurred during the period: the Pliensbachian-Toarcian extinction in the Early Jurassic, and the end Jurassic transition, which is disputed in its impact.[4]
The Jurassic period is divided into three epochs: Early, Middle, and Late. Similarly, in stratigraphy, the Jurassic is divided into the Lower Jurassic, Middle Jurassic, and Upper Jurassic series of rock formations.
By the beginning of the Jurassic, the supercontinent Pangaea had begun rifting into two landmasses: Laurasia to the north, and Gondwana to the south. This created more coastlines and shifted the continental climate from dry to humid, and many of the arid deserts of the Triassic were replaced by lush rainforests.
On land, the fauna transitioned from the Triassic fauna, dominated by both dinosauromorph and pseudosuchian archosaurs, to one dominated by dinosaurs alone. The first birds also appeared during the Jurassic, having evolved from a branch of theropod dinosaurs. Other major events include the appearance of the earliest lizards, and the evolution of therian mammals. Crocodilians made the transition from a terrestrial to an aquatic mode of life. The oceans were inhabited by marine reptiles such as ichthyosaurs and plesiosaurs, while pterosaurs were the dominant flying vertebrates.
Etymology and history
The chronostratigraphic term "Jurassic" is directly linked to the Jura Mountains, a mountain range mainly following the course of the France–Switzerland border. The name "Jura" is derived from the Celtic root *jor via Gaulish *iuris "wooded mountain", which, borrowed into Latin as a place name, evolved into Juria and finally Jura.[5][6][7] During a tour of the region in 1795,[note 2] Alexander von Humboldt recognized the mainly limestone dominated mountain range of the Jura Mountains as a separate formation that had not been included in the established stratigraphic system defined by Abraham Gottlob Werner, and he named it "Jura-Kalkstein" ('Jura limestone') in 1799.[note 3][5][6][10]
Thirty years later, in 1829, the French naturalist Alexandre Brongniart published a survey on the different terrains that constitute the crust of the Earth. In this book, Brongniart referred to the terrains of the Jura Mountains as terrains jurassiques, thus coining and publishing the term for the first time.[11] The German geologist Leopold von Buch in 1839 established the three-fold division of the Jurassic, originally named from oldest to youngest, the Black Jurassic, Brown Jurassic and White Jurassic.[12] The term "Lias" had previously been used equivalently for strata of equivalent age to the Black Jurassic in England by Conybeare and Phillips in 1822. French palaeontologist Alcide d'Orbigny in papers between 1842 and 1852 would divide the Jurassic into ten stages “étages” based on ammonite and other fossil assemblages in England and France, of which seven are still used, though none retain the original definition. German geologist and palaeontologist Friedrich August von Quenstedt in 1858 would divide the three series of von Buch in the Swabian Jura into six subdivisions defined by ammonites and other fossils. German palaeontologist Albert Oppel in studies between 1856 and 1858 altered d'Orbigny's original scheme and further subdivided the stages into biostratigraphic zones, based primarily on ammonites. Most of the modern stages of the Jurassic were formalized at the "Colloque du Jurassique á Luxembourg" in 1962.[13]
Geology
The Jurassic period is divided into three epochs: Early, Middle, and Late. Similarly, in stratigraphy, the Jurassic is divided into the Lower Jurassic, Middle Jurassic, and Upper Jurassic series of rock formations, also known in Europe as Lias, Dogger and Malm.[14] The three epochs are subdivided into shorter spans of time called ages. The ages of the Jurassic from youngest to oldest are:
| Upper/Late Jurassic | Tithonian | (152.1 ± 4 – 145 ± 4 Mya) |
| Kimmeridgian | (157.3 ± 4 – 152.1 ± 4 Mya) | |
| Oxfordian | (163.5 ± 4 – 157.3 ± 4 Mya) | |
| Middle Jurassic | Callovian | (166.1 ± 4 – 163.5 ± 4 Mya) |
| Bathonian | (168.3 ± 3.5 – 166.1 ± 4 Mya) | |
| Bajocian | (170.3 ± 3 – 168.3 ± 3.5 Mya) | |
| Aalenian | (174.1 ± 2 – 170.3 ± 3 Mya) | |
| Lower/Early Jurassic | Toarcian | (182.7 ± 1.5 – 174.1 ± 2 Mya) |
| Pliensbachian | (190.8 ± 1.5 – 182.7 ± 1.5 Mya) | |
| Sinemurian | (199.3 ± 1 – 190.8 ± 1.5 Mya) | |
| Hettangian | (201.3 ± 0.6 – 199.3 ± 1 Mya) |
Stratigraphy
Jurassic stratigraphy is primarily based around of the use of ammonites as index fossils, with the First Appearance Datum of specific ammonite taxa being used to mark the beginnings of stages, and well as smaller timespans within stages, referred to as "Ammonite Zones", these in turn are also sometimes subdivided further into subzones. Global stratigraphy is based on standard European ammonite zones, with other regions being calibrated to the European successions.[13]

The oldest part of the Jurassic period has historically been referred to as the Lias or Liassic, roughly equivalent in extent to the Early Jurassic, but also including part of the preceding Rhaetian. The Hettangian stage was named by Swiss palaeontologist Eugène Renevier in 1864 after Hettange-Grande in North-Eastern France. The Global Boundary Stratotype Section and Point (GSSP) for the base of the Hettangian is located at Kuhjoch pass, Karwendel Mountains, Northern Calcareous Alps, Austria, which was ratified in 2010. The beginning of the Hettangian, and thus the Jurassic as a whole, is marked by the first appearance of the ammonite Psiloceras spelae tirolicum in the Kendlbach Formation exposed at Kuhjoch.[15] The base of the Jurassic was previously defined as the first appearance of Psiloceras planorbis by Albert Oppel in 1856–58, but this was changed as the appearance was seen as too localised an event for an international boundary.[13]
The Sinemurian stage was defined and introduced into scientific literature by Alcide d'Orbigny in 1842. It takes its name from the French town of Semur-en-Auxois, near Dijon. The original definition of Sinemurian included what is now the Hettangian. The GSSP of the Sinemurian is located at a cliff face north of the hamlet of East Quantoxhead, 6 kilometres east of Watchet, Somerset, England, within the Blue Lias. The beginning of the Sinemurian is defined by the first appearance of the ammonite Vermiceras quantoxense.[13][16]
The Pliensbachian was named by German palaeontologist Albert Oppel in 1858 after the hamlet of Pliensbach in the community of Zell unter Aichelberg in the Swabian Alb, near Stuttgart, Germany. The GSSP for the base of the Pliensbachian is found at the Wine Haven locality in Robin Hood's Bay, Yorkshire, England, in the Redcar Mudstone Formation. The beginning of the Pliensbachian is defined by the first appearance of the ammonite Bifericeras donovani.[17]
The Toarcian is named after the village Thouars (Latin: Toarcium), just south of Saumur in the Loire Valley of France, it was defined by Alcide d'Orbigny in 1842 originally from Vrines quarry around 2 km northwest of the village. The GSSP for the base of the Toarcian is located at Peniche, Portugal. The boundary is defined by the first appearance of ammonites belonging to the subgenus Dactylioceras (Eodactylioceras).[18]
The Aalenian is named after the city of Aalen in Germany. The Aalenian was defined by Swiss geologist Karl Mayer-Eymar in 1864. The lower boundary was originally between the dark clays of the Black Jurassic and the overlying clayey sandstone and ferruginous oolite of the Brown Jurassic sequences of southwestern Germany.[13] The GSSP for the base of the Aalenian is located at Fuentelsaz in the Iberian range near Guadalajara, Spain. The base of the Aalenian is defined by the first appearance of the ammonite Leioceras opalinum.[19]
The Bajocian is named after the town of Bayeux (Latin: Bajoce) in Normandy, France, and was defined by Alcide d'Orbigny in 1842. The GSSP for the base of the Bajocian is located at Murtinheira in Portugal, and was defined in 1997. The base of the Bajocian is defined by the first appearance of the ammonite Hyperlioceras mundum.[20]
The Bathonian is named after the city of Bath, England, introduced by Belgian geologist d'Omalius d'Halloy in 1843, after an incomplete section of oolitic limestones in several quarries in the region. The GSSP for the base of the Bathonian is Ravin du Bès, Bas-Auran area, Alpes de Haute Provence, France, which was defined in 2009. The base of the Bathonian is defined by the first appearance of the ammonite Gonolkites convergens, at the base of the Zigzagiceras zigzag ammonite zone.[21]
The Callovian is derived from the Latinized name of the village of Kellaways in Wiltshire, England, and was defined by Alcide d'Orbigny in 1852, originally with base at the contact between the Forest Marble Formation and the Cornbrash Formation. However, this boundary was later found to be situated within the upper part of the Bathonian. The base of the Callovian does not yet have a certified GSSP, as of 2019.[13]
The Oxfordian is named after the city of Oxford in England, and was named by Alcide d'Orbigny in 1844 in reference to the Oxford Clay. The base of the Oxfordian lacks a defined GSSP. W. J. Arkell in studies in 1939 and 1946 placed the lower boundary of the Oxfordian as the first appearance of the ammonite Quenstedtoceras mariae (then placed in the genus Vertumniceras). Subsequent proposals have suggested the first appearance of Cardioceras redcliffense as the lower boundary.[13]
The Kimmeridgian is named after the village of Kimmeridge on the coast of Dorset, England. It was named by Alcide d'Orbigny in 1842, in reference to the Kimmeridge Clay. Although not confirmed, the Flodigarry section at Staffin Bay on the Isle of Skye, Scotland has been submitted as the GSSP for the base of the Kimmeridgian.[22]
The Tithonian was introduced in scientific literature by Albert Oppel in 1865. The name Tithonian is unusual in geological stage names because it is derived from Greek mythology rather than a placename. Tithonus was the son of Laomedon of Troy and fell in love with Eos, the Greek goddess of dawn. His name was chosen by Albert Oppel for this stratigraphical stage because the Tithonian finds itself hand in hand with the dawn of the Cretaceous. The base of the Tithonian currently lacks a GSSP.[13] The upper boundary of the Jurassic is also currently undefined. Calpionellids, an enigmatic group of pelagic protists with urn shaped calcitic tests briefly abundant during the latest Jurassic to earliest Cretaceous, have been suggested to represent the most promising candidates for fixing the J/K boundary.[23]
Mineral and hydrocarbon deposits
The Kimmeridge Clay and equivalents are the major source rock for the North Sea oil.[24] The Arabian Intrashelf Basin, deposited from the late Middle to Upper Jurassic, is the setting of the world's largest oil reserves, including the Ghawar Field, the world largest oil field.[25] The Jurassic aged Sargelu[26] and Naokelekan Formations[27] are major source rocks for oil in Iraq. Over 1500 gigatons of Jurassic coal reserves are found in North-West China, primarily in the Turpan-Hami Basin and the Ordos Basin.[28]
Impact craters
Major impact craters include the Morokweng crater, a 70 km diameter crater buried beneath the Kalahari desert in northern South Africa. The impact is dated to the Jurassic-Cretaceous boundary, around 145 Ma. The Morokweng crater has been suggested to have had a role in the turnover at the Jurassic-Cretaceous transition.[29] Another major impact crater is the Puchezh-Katunki crater, 40-80 kilometres in diameter, buried beneath Nizhny Novgorod Oblast, Russia. The impact has been dated to the Sinemurian, around 192-196 Mya.[30]
Paleogeography and tectonics
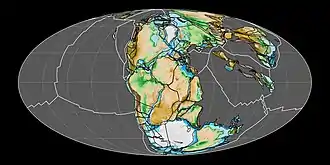

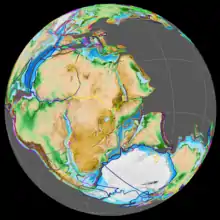
During the early Jurassic period, the supercontinent Pangaea broke up into the northern supercontinent Laurasia and the southern supercontinent Gondwana; the Gulf of Mexico opened in the new rift between North America and what is now Mexico's Yucatán Peninsula. The Jurassic North Atlantic Ocean was relatively narrow, while the South Atlantic did not open until the following Cretaceous period.[31] The continents were surrounded by Panthalassa, with the Tethys Ocean between Gondwana and Asia. At the end of the Triassic, there was a marine transgression in Europe, flooding most parts of central and western Europe transforming it into an archipelago of islands surrounded by shallow seas.[32] The Boreal Ocean was connected to the western Tethys by the "Viking corridor", a several hundred kilometer wide passage between the Baltic Shield and Greenland.[33] Madagascar and Antarctica began to rift away from Africa during Early Jurassic, beginning the fragmentation of Gondwana.[34][35] At the beginning of the Jurassic, North and South America remained connected, but by the beginning of the Late Jurassic had rifted apart, forming the Caribbean Seaway, connecting the western Tethys with eastern Panthalassa.[36] During the Early Jurassic, around 190 million years ago, the Pacific Plate originated at the triple junction of the Farallon, Phoenix, and Izanagi plates, the three main oceanic plates of Panthalassa. The previously stable triple junction had converted to an unstable arrangement surrounded on all sides by transform faults, due to a kink in one of the plate boundaries, resulting in the formation of the Pacific Plate at the centre of the junction, which began to expand.[37] During the Middle to early Late Jurassic, the Sundance Seaway, a shallow epicontinental sea would cover much of northwest North America.[38]
Based on estimated sea level curves, the eustatic sea level was close to present levels during the Hettangian and Sinemurian, rising several tens of metres during the late Sinemurian-Pliensbachian, before regressing to near present levels by the late Pliensbachian. There seems to have been a gradual rise to a peak of ~75 m above present sea level during the Toarcian. During the latest part of the Toacian, the sea level again drops by several tens of metres. The sea level progressively rose from the Aalenian onwards, aside from dips of a few tens of metres in the Bajocian and around the Callovian-Oxfordian boundary, culminating in a sea level possibly as high as 140 metres above present sea level at the Kimmeridgian-Tithonian boundary. The sea levels falls in the Late Tithonian, perhaps to around 100 metres, before rebounding to around 110 metres at the Tithonian-Berriasian boundary. Sea level within the long-term trend was cyclical with 64 fluctuations through the Jurassic, 15 of which were over 75 metres. The most noted cyclicity in Jurassic rocks is fourth order, with a periodicity of approximately 410,000 years.[39]
The Jurassic was a time of calcite sea geochemistry in which low-magnesium calcite was the primary inorganic marine precipitate of calcium carbonate. Carbonate hardgrounds were thus very common, along with calcitic ooids, calcitic cements, and invertebrate faunas with dominantly calcitic skeletons.[40] The rise of calcareous plankton during the Middle Jurassic profoundly altered ocean chemistry, with the deposition of biomineralizing plankton on the ocean floor acting as a buffer against large CO2 emissions.[41]
The first of several massive batholiths were emplaced in the northern American cordillera beginning in the mid-Jurassic, marking the Nevadan orogeny.[42]
In Africa, Early Jurassic strata are distributed in a similar fashion to Late Triassic beds, with more common outcrops in the south and less common fossil beds which are predominated by tracks to the north.[43] As the Jurassic proceeded, larger and more iconic groups of dinosaurs like sauropods and ornithopods proliferated in Africa.[43] Middle Jurassic strata are neither well represented nor well studied in Africa.[43] Late Jurassic strata are also poorly represented apart from the spectacular Tendaguru fauna in Tanzania.[43] The Late Jurassic life of Tendaguru is very similar to that found in western North America's Morrison Formation.[43]
 Jurassic limestones and marls (the Matmor Formation) in southern Israel
Jurassic limestones and marls (the Matmor Formation) in southern Israel The Upper Jurassic Morrison Formation in Colorado is one of the most fertile sources of dinosaur fossils in North America
The Upper Jurassic Morrison Formation in Colorado is one of the most fertile sources of dinosaur fossils in North America Gigandipus, a dinosaur footprint in the Lower Jurassic Moenave Formation at the St. George Dinosaur Discovery Site at Johnson Farm, southwestern Utah
Gigandipus, a dinosaur footprint in the Lower Jurassic Moenave Formation at the St. George Dinosaur Discovery Site at Johnson Farm, southwestern Utah The Permian through Jurassic stratigraphy of the Colorado Plateau area of southeastern Utah
The Permian through Jurassic stratigraphy of the Colorado Plateau area of southeastern Utah
Climate
The climate of Jurassic was generally warmer than at present, by around 5 °C to 10 °C. Atmospheric carbon dioxide levels were likely four times higher than present. Forests likely grew near the poles, and experienced warm summers and cold, sometimes snowy winters, and there were unlikely to have been ice sheets given the high summer temperatures, though mountain glaciers may have existed. The ocean depths were likely 8 °C warmer than present, and reefs grew 10° of latitude further north and south. The Intertropical Convergence Zone likely existed over the oceans, resulting in large areas of desert in the lower latitudes.[44]
Toarcian Oceanic Anoxic Event
The Toarcian Oceanic Anoxic Event (TOAE) was an episode of widespread oceanic anoxia during the early part of the Toarcian period, c. 183 Mya. It is marked by a globally documented high amplitude negative carbon isotope excursion,[45] as well as the deposition of black shales, and the extinction and collapse of carbonate producing marine organisms. The cause is often linked to the eruption of the Karoo-Ferrar large igneous provinces and the associated increase of carbon dioxide concentration in the atmosphere and the possible associated release of methane clathrates. This likely accelerated the hydrological cycle and increased silicate weathering. Groups affected include ammonites, ostracods, foraminifera, brachiopods, bivalves and cnidarians,[46][47] with the last two spire-bearing brachiopod orders Spiriferinida and Athyridida becoming extinct.[48] While the event had significant impact on marine invertebrates, it had little effect on marine reptiles.[49] During the TOAE, the Sichuan Basin was transformed into a giant lake, probably 3 times the size of Lake Superior, represented by the Da’anzhai Member of the Ziliujing Formation. The lake likely sequestered ∼460 Gigatons (Gt) of organic carbon and ∼1,200 Gt of inorganic carbon during the event.[50] During the event. seawater PH, which had already substantially decreased prior to the event, increased slightly during the early stages of the TOAE, before dropping to its lowest point around the middle of the event.[51] This ocean acidification is what likely caused the collapse of carbonate production.[52][53]
End-Jurassic transition
The end-Jurassic transition was originally considered one of eight mass extinctions, but is now considered to be complex interval of faunal turnover, with the increase in diversity of some groups and decline in others, though the evidence for this is primarily European, probably controlled by changes in eustatic sea level.[29]
Flora
End-Triassic extinction
The preceding end-Triassic extinction would result in the decline of Peltaspermaceae seed ferns, with Lepidopteris perisisting into the Early Jurassic in Patagonia.[54] At the Triassic-Jurassic boundary in Greenland, the sporomorph diversity suggests a complete floral turnover.[55] An analysis of macrofossil floral communities in Europe suggests no extinction over the Triassic-Jurassic boundary, and that changes were mainly due to local ecological succession.[56] Dicroidium, a seed fern that was a dominant part of Gondwanan floral communities during the Triassic, would decline at the T-J, boundary, surviving as a relict in Antarctica into the Sinemurian.[57]
Floral composition
Flowering plants, which make up 90% of living plant species, have no records from the Jurassic, with no claim of Jurassic representatives of the group having gained widespread acceptance.[58]
Conifers

Trees of the Jurassic were dominated by conifers and modern conifier groups would diversify throughout the period.
Araucarian conifers were widespread across both hemispheres. The divergence between Araucaria and the branch containing Wollemia and Agathis is estimated to have taken place during the Mid-Jurassic, based on Araucaria mirabilis and Araucaria sphaerocarpa from the Middle Jurassic of Argentina and England respectively, which are early members of the Araucaria lineage. Representatives of Wollemia-Agathis lineage are not known until the Cretaceous.[59][60]
Also abundant during the Jurassic is the extinct family Cheirolepidiaceae, often recognised by their highly distinctive Classopolis pollen. Jurassic representatives include the pollen cone Classostrobus and the seed cone Pararaucaria. Both Araucarian and Cheirolepidiaceae confiers often occur in association.[61]
The oldest definitive record of the cypress family (Cupressaceae) is Austrohamia minuta from the Early Jurassic (Pliensbachian) of Patagonia, known from several elements.[62] Austrohamia is thought to have close affinities with Taiwania and Cunninghamia. By the Mid-Late Jurassic Cupressaceae were abundant in warm temperate-tropical regions of the Northern Hemisphere, most abundantly represented by the genus Elatides.[63] The seed cone Scitistrobus from the Middle Jurassic (Aalenian) of Scotland displays a mosaic of traits indicative of ancestral Voltziales and derived Cupressaceae.[64]
The oldest record of the pine family (Pinaceae) is the seed cone Eathiestrobus, known from the Late Jurassic (Kimmeridgian) also of Scotland.[65] During the Early Jurassic, the flora of the mid-latitudes of Eastern Asia were dominated by the extinct deciduous broad leafed conifer Podozamites, likely of voltzialean affinities, with its range extending northwards into polar latitudes of Siberia, but its range contracted northward in the Middle-Late Jurassic corresponding to the increasing aridity of the region.[66]
The earliest record of the yew family (Taxaceae) is Palaeotaxus rediviva, from the Hettangian of Sweden, suggested to be closely related to Austrotaxus, while Marskea jurassica from the Middle Jurassic of Yorkshire, England and material from the Callovian-Oxfordian Daohugou bed in China are thought to be closely related to Amentotaxus. The Daohugou material in particular is extremely similar to living Amentotaxus, only differing in having shorter seed-bearing axes.[67]
Podocarpaceae, today largely confined to the Southern Hemisphere occurs in the Northern Hemisphere during the Jurassic, including Podocarpophyllum from the Lower-Middle Jurassic of Central Asia and Siberia.[68] Scarburgia from the Middle Jurassic of Yorkshire,[69] and Harrisiocarpus from the Jurassic of Poland.[70]
Ginkgoales
Ginkgoales, which are currently represented by the single living species Ginkgo biloba, were more diverse during the Jurassic, they were among the most important components of Laurasian Jurassic floras, and were adapted to a wide variety of climatic conditions. Based on reproductive organs several lineages can be distinguished, including Yimaia, Grenana, Nagrenia and Karkenia, alongside Ginkgo. These lineages are associated with leaf morphotaxa such as Baiera, Ginkgoites and Sphenobaiera, some of which overlap with the morphological variability and growth stages of living Ginkgo biloba leaves and therefore cannot be used for reliable taxonomic identification.[71][72] Umaltolepis, historically thought to be ginkgoalean, and Vladimaria from the Jurassic of Asia have strap shaped ginkgo-like leaves (Pseudotorellia), with highly distinct reproductive structures with similarities to those of peltasperm and corystosperm seed ferns, and have been placed in the separate order Vladimariales, which may belong to a broader Ginkgoopsida.[73]
Ferns
The ground cover was dominated by ferns, including members of the living families Dipteridaceae, Matoniaceae, Osmundaceae and Marattiaceae,[74] as well as horsetails. Polypodiales, which today make up 80% of living fern diversity, have no record from the Jurassic, and are thought to have diversified in the Cretaceous,[75] though the widespread Jurassic herbaceous fern genus Coniopteris, historically interpreted as a close relative of tree ferns of the family Dicksoniaceae, has recently been reinterpreted as an early relative of the group.[76]
A calicfied rhizome of an Osmundaceous fern from the Early Jurassic of Sweden belongs to the stem group of the living genus Osmundastrum, with the preservation showing the remains of chromosomes during cell division.[77] An analysis of the Osmundastrum rhizome found that it had been interacted with by numerous organisms, including lycopsid roots growing into the rhizome, probable peronosporomycetes as well as boring and coprolites likely by orbatid mites.[78]
The oldest remains of modern horsetails of the genus Equisetum first appear in the Early Jurassic, represented by Equisetum dimorphum from the Early Jurassic of Patagonia[79] and Equisetum laterale from the Early-Middle Jurassic of Australia.[80][81] Silicified remains of Equisetum thermale from the Late Jurassic of Argentina exhibit all the morphological characters of modern members of the genus.[82] The estimated split between Equisetum bogotense and all other living Equisetum is estimated to have occurred no later than the Early Jurassic.[81]
The Cyatheales, the group containing most modern tree ferns would appear during the Late Jurassic, represented by members of the genus Cyathocaulis, which are suggested to be early members of Cyatheaceae based on cladistic analysis.[83] Only a handful of possible records exist of the Hymenophyllaceae are known from the Jurassic, including Hymenophyllites macrosporangiatus from the Russian Jurassic.[84]
Bennettitales
Bennettitales are a group of seed plants widespread throughout the Mesozoic with foliage bearing strong similarities to those of cycads, to the point of morphologically indistinguishable. Benettitales can be distinguished from cycads by the fact they have a different arrangement of stomata, and are not thought to be closely related.[85] Benettitales have morphologies varying from cycad-like to shubs and small trees. The Williamsoniaceae grouping is thought to have had a divaricate branching habit, similar to living Banksia, and adapted to growing in open habitats with poor soil nutrient conditions.[86] Benettitales exhibit complex, flower like reproductive structures that are thought to have been pollinated by insects. Several groups of insects that bear long proboscis, including extinct families like Kalligrammatid lacewings[87] and extant Acroceridae flies,[88] are suggested to have been pollinators of benettitales, feeding on nectar produced by bennettitalean cones.
Cycads
Cycads were present during the Jurassic, the living groups of cycads have been suggested to have diverged from each other in the Early Jurassic,[89] though a later analysis placed this divergence during the Late Permian, which placed the diversification of the Zamiineae cycads during the Jurassic.[90] Cycads are difficult to distinguish from Bennettitales based on leaf morphology alone. Cycads are thought to have been a relatively minor component of mid-Mesozoic floras.[91] and mostly confined to tropical and subtropical latitudes.[92] Cycad foliage is assigned to morphogenera including Ctenis and Pterophyllum, but are not phylogenetically informative. Seeds from the late Callovian-early Oxfordian Oxford Clay are definitively assignable to the living family Cycadaceae.[93] While seeds found in the gut of the dinosaur Isaberrysaura from the Middle Jurassic of Argentina are assigned to Zamiineae, which includes all other living cycads.[94] The Nilssoniales, such as the leaf genus Nilssonia with leaves morphologically similar to those of cycads, have often been considered cycads or cycad relatives, but have been found to be distinct, perhaps more closely allied with Bennettitales.[92]
Gnetophytes
Protognetum from the Middle Jurassic of China is oldest known member of the gnetophytes and the only one known from the Jurassic. It exhibits characteristics of both Gnetum and Ephedra, and is placed in the monotypic family Protognetaceae.[95]
Seed ferns

Seed ferns (Pteridospermatophyta) is a collective term to refer to disparate lineages of fern like plants that produce seeds, with uncertain affinities to living seed plant groups. Prominent groups of Jurassic seed ferns include Caytoniales, which includes the leaf taxon Sagenopteris, Caytonanthus pollen structures and Caytonia ovulate structures, often found in close association. They have frequently been suggested to have been closely related or perhaps ancestral to flowering plants, but no definitive evidence of this has been discovered.[96] The other prominent group is the Corystospermales, including genera like the leaf genus Pachypteris, prominent in the Jurassic of the Northern Hemisphere. As well as pollen organs belonging to Pteruchus and Umkomasia ovulate structures.[97][98]
Czekanowskiales
Czekanowskiales, also known as Leptostrobales, are a group of gymnosperms of uncertain affinities with persistent leaves borne on deciduous short shoots, subtended by scale-like leaves, known from the Late Triassic (possibly Late Permian[99]) to Cretaceous.[100] They are thought to have had a tree or shrub like habit, and formed a conspicuous component of Mesozoic temperate and warm–temperate floras.[99] Jurassic genera include the leaf genera Czekanowskia, Phoenicopsis and Solenites, associated with the ovulate cone Leptostrobus.[100]
Pentoxylales
The Pentoxylales are a small group of gymnosperms of obscure affinities, known from the Jurassic and Cretaceous of Gondwana. These include the stems Pentoxylon, strap-shaped leaves Taeniopteris (more broadly used as a morphogenus representing other plant types) and Nipaniophyllum for well preserved leaves, Sahnia pollen organs, and Carnoconites seed-bearing structures. The habit of the group is uncertain, but may have been small trees.[100]
Quillworts
Quillworts virtually identical to modern species are known from the Jurassic onwards. Isoetites rolandii from the Middle Jurassic of Oregon is the earliest known species to represent all major morphological features of modern Isoetes.[101]
Moss
The moss Kulindobryum from the Middle Jurassic of Russia is thought to have affinites with the Splachnaceae, while Bryokhutuliinia from the same region is thought to have affinities with Dicranales.[102] Heinrichsiella from the Jurassic of Patagonia is thought to belong to the families Polytrichaceae or Timmiellaceae basal to Bryidae, and is the oldest representative of the grade.[103]
Liverworts
The liverwort Pellites hamiensis from the Middle Jurassic Xishanyao Formation of China is the oldest record of the family Pelliaceae.[104] Pallaviciniites sandaolingensis from the same deposit is thought to belong to the subclass Pallaviciniineae within the Pallaviciniales.[105] Ricciopsis sandaolingensis also from the same deposit is the only Jurassic record of Ricciaceae.[106]
Regional abundance
In an analysis of the ferns of the Hettangian aged Mecsek Coal Formation found that the predominant groups of ferns by order of abundance belonged to the families Dipteridaceae (48% of collected specimens) Matoniaceae (25%), Osmundaceae (21%), Marattiaceae (6%) and 3 specimens of Coniopteris. They found that most of the ferns likely grew in monospecific thickets in disturbed areas.[74] The Middle-Late Jurassic Daohugou flora of China was dominated by Gymnosperms and ferns, with the most abundant group of gymnosperms being Bennettitales, followed by conifers and ginkgophytes.[107] High latitude floras of the New Zealand Jurassic were of low diversity, with only 43 species being recorded dominated by "conifers, ferns, bennettitaleans, pentoxylaleans and locally, equisetaleans" with Ginkgoales being entirely absent.[108] The flora of the Middle Jurassic Stonesfield Slate of England was dominated by "araucariacean and cheirolepidiacean conifers, bennettitaleans, and leaves of the possible gymnosperm Pelourdea" representing a coastal environment.[109]
Fauna
Aquatic and marine
During the Jurassic period, the primary vertebrates living in the sea were fish and marine reptiles. The latter include ichthyosaurs, which were at the peak of their diversity, plesiosaurs, including pliosaurs, and marine thalattosuchian crocodyliformes of the families Teleosauridae, Machimosauridae and Metriorhynchidae.[110]
Calcareous sabellids (Glomerula) appeared in the Early Jurassic.[111][112] The Jurassic also had diverse encrusting and boring (sclerobiont) communities, and it saw a significant rise in the bioerosion of carbonate shells and hardgrounds. Especially common is the ichnogenus (trace fossil) Gastrochaenolites.[113] During the Jurassic period, about four or five of the twelve clades of planktonic organisms that exist in the fossil record either experienced a massive evolutionary radiation or appeared for the first time.[14]
 Ichthyosaurus from lower (early) Jurassic slates in southern Germany featured a dolphin-like body shape.
Ichthyosaurus from lower (early) Jurassic slates in southern Germany featured a dolphin-like body shape. Plesiosaurs like Muraenosaurus roamed Jurassic oceans.
Plesiosaurs like Muraenosaurus roamed Jurassic oceans.
Terrestrial

On land, various archosaurian reptiles remained dominant. The Jurassic was a golden age for the large herbivorous dinosaurs known as the sauropods—Camarasaurus, Apatosaurus, Diplodocus, Brachiosaurus, and many others—that roamed the land late in the period; their foraging grounds were either the prairies of ferns, palm-like cycads and bennettitales, or the higher coniferous growth, according to their adaptations. The smaller Ornithischian herbivore dinosaurs, like stegosaurs and small ornithopods were less predominant, but played important roles. They were preyed upon by large theropods, such as Ceratosaurus, Megalosaurus, Torvosaurus and Allosaurus, all these belong to the 'lizard hipped' or saurischian branch of the dinosaurs.[114]
During the Late Jurassic, the first avialans, like Archaeopteryx, evolved from small coelurosaurian dinosaurs. In the air, pterosaurs were common; they ruled the skies, filling many ecological roles now taken by birds,[115] and may have already produced some of the largest flying animals of all time.[116][117] Within the undergrowth were various types of early mammals, as well as tritylodonts, lizard-like sphenodonts, and early lissamphibians. The rest of the Lissamphibia evolved in this period, introducing the first salamanders and caecilians.[118]
 Diplodocus, reaching lengths over 30 m, was a common sauropod during the late Jurassic.
Diplodocus, reaching lengths over 30 m, was a common sauropod during the late Jurassic. Allosaurus was one of the largest land predators of the Jurassic.
Allosaurus was one of the largest land predators of the Jurassic.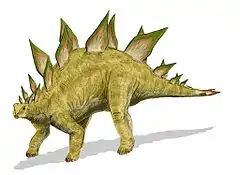 Stegosaurus is one of the most recognizable genera of dinosaurs and lived during the mid to late Jurassic.
Stegosaurus is one of the most recognizable genera of dinosaurs and lived during the mid to late Jurassic. Aurornis xui, which lived in the late Jurassic, may be the most primitive avialan dinosaur known to date, and is one of the earliest avialans found to date.
Aurornis xui, which lived in the late Jurassic, may be the most primitive avialan dinosaur known to date, and is one of the earliest avialans found to date.
Crocodylomorphs


The T-J extinction decimated pseudosuchian diversity, with crocodylomorphs, which originated during the early Late Triassic (Norian), being the only group of pseudosuchians to survive the T-J extinction. The morphological diversity of crocodylomorphs during the Early Jurassic was around the same as those of Late Triassic pseudosuchians, but occupied different areas of morphospace, suggesting that they occupied different ecological niches to their Triassic counterparts, and that there was an extensive and rapid radiation of crocodylomorphs during this interval.[119] While living crocodilians are confined to an aquatic ambush predator lifestyle, Jurassic crocodyliformes exhibit a wide variety of life habits, with the earliest known herbivorous crocodylomorph being an unnamed species of protosuchid from the Early Jurassic of Arizona.[120] The earliest lineages of Crocodylomorpha are represented by the terrestrial small-bodied long-legged "sphenosuchians", one lineage of which, the Hallopodidae, perisisted into the Late Jurassic.[121][122] The Thalattosuchia, a clade of predominantly marine crocodylomorphs including both teleosaurioids and metriorhynchoids, first appeared during the Early Jurassic (Toarcian), and became prominent part of marine reptile communities during the Jurassic. During the Jurassic, the Neosuchia would appear, the Neosuchia are thought to be ancestrally freshwater aquatic, similar to living crocodilians.[123] The oldest record of Neosuchians has been suggested to be Calsoyasuchus, from the Early Jurassic of Arizona, which in many analyses has been recovered as the earliest branching goniopholidid, which radically alters times of diversification for crocodylomorphs, however, this placement has been disputed, with some analyses finding it outside Neosuchia, which would place the oldest records of Neosuchia in the Middle Jurassic.[123] Razanandrongobe from the Middle Jurassic of Madagascar has been suggested the represent the oldest record of Notosuchia, a primarily Gondwanan clade of mostly terrestrial crocodylomorphs, otherwise known from the Cretaceous.[124]
Turtles
Stem-group turtles (Testudinata) would diversify during the Jurassic. Jurassic stem-turtles belong to two progressively more advanced clades, the Mesochelydia and Perichelydia.[125] It is thought that the ancestral condition for Mesochelydians is aquatic, as opposed to terrestrial for Testudinata.[126] The two modern groups of turtles (Testudines), Pleurodires and Cryptodires, would diverge by the Middle Jurassic.[127][128] Early cryptodire lineages like Xinjiangchelyidae are known from the Middle Jurassic,[129] while an early stem pleurodire lineage, the ocean-going Platychelyidae is known from the Late Jurassic.[130] The Thalassochelydia, a diverse lineage of sea-turtles, is known from the Late Jurassic (Oxfordian-Tithonian) of Europe and South America.[131]
Lepidosaurs
Lepidosaurs, which include squamates (lizards and snakes) and rhynchocephalians (which today only includes the tuatara) would diversify during the Jurassic, however little can be said for diversity trends due to the poor fossil record during this interval.[132] Rynchocephalians reached their highest morphological diversity in their evolutionary history during the Jurassic, occupying a wide range of lifestyles, including aquatic taxa such as pleurosaurids, the specialized herbivorous "eilenodontines" as well as Oenosaurus, which had broad tooth plates indicative of durophagy. Cynosphenodon from the Early Jurassic of Mexico is suggested to be among the closest known relatives of the living tuatara.[133] Rhynchocephalians disappeared from Asia after the Early Jurassic.[134] Crown group squamates first appeared during the Middle Jurassic of Asia and Europe,[132] with the estimated origin of living squamates during the Early Jurassic (~190 Mya) and the divergence of most major squamate groups during the Early-Middle Jurassic.[135] Many Jurassic squamates have unclear relationships to living groups.[136] Early members of the snake lineage (Ophidia) appear during the Middle Jurassic,[137] as well as the Paramacellodidae, a family of lizards which are diagnosed by their distinctive covering of osteoderms, and are thought to be early members of Scincomorpha.[138] Eichstaettisaurus from the Late Jurassic of Germany has been suggested to be an early relative of geckos, and displays adaptations for climbing.[139] Dorsetisaurus from the Late Jurassic of North America and Europe represents the oldest widely accepted record of Anguimorpha.[140] Marmoretta from the Middle Jurassic of Britain represents a late surviving lepidosauromorph outside both groups.[141]
.jpg.webp) Vadasaurus herzogi, a rynchocephalian from the Upper Jurassic Solnhofen Limestone of Germany
Vadasaurus herzogi, a rynchocephalian from the Upper Jurassic Solnhofen Limestone of Germany Homeosaurus maximiliani a rynchocephalian from the Solnhofen Limestone
Homeosaurus maximiliani a rynchocephalian from the Solnhofen Limestone Pleurosaurus, an aquatic rhynchocephalian from the Late Jurassic of Europe
Pleurosaurus, an aquatic rhynchocephalian from the Late Jurassic of Europe Eichstaettisaurus schroederi, an extinct lizard from the Solnhofen Limestone.
Eichstaettisaurus schroederi, an extinct lizard from the Solnhofen Limestone.
Choristoderes
The earliest known remains of Choristodera, a group of freshwater aquatic reptiles with uncertain affinities to other reptile groups, are found in the Middle Jurassic. Only two genera of choristodere are known from the Jurassic. One is Cteniogenys, thought to be the most basal known choristodere, which is known from the Middle-Late Jurassic of Europe and Late Jurassic of North America, with similar remains also known from the upper Middle Jurassic of Kyrgyzstan, European Russia and western Siberia.[142] The other is Coeruleodraco from the Late Jurassic of China, which has close affinities to a clade of non-Neochoristodera from the Early Cretaceous of Asia as well as Lazarussuchus from the Cenozoic of Europe.[143]
Ichthyosaurs

Ichthyosaurs suffered an evolutionary bottleneck, with all non-neoichthyosaurians becoming extinct at the end of the Triassic. Ichthyosaurs would reach their apex of species diversity during the Early Jurassic, with an array of morphologies including the huge apex predator Temnodontosaurus and swordfish-like Eurhinosaurus. Jurassic ichthyosaurs were significantly less morphologically diverse than their Triassic counterparts, and would become less diverse and morphologically conservative after the Early Jurassic.[144] The fossil record of ichthyosaurs during the Middle Jurassic is poor.[145] The Ophthalmosauridae, which would represent all (with the exception of Malawania) ichthyosaurs from the late Middle Jurassic onwards, first appear in the early Middle Jurassic (Bajocian), before spreading worldwide by the Late Jurassic.[146]
Plesiosaurs

Plesiosaurs originated at the end of the Triassic (Rhaetian). By the Triassic boundary, all other sauropterygians, including placodonts and nothosaurs had become extinct. At least six lineages of plesiosaur crossed the T-J boundary.[147] Plesiosaurs were already diverse in the earliest Jurassic, with the majority of plesiosaurs in the Hettangian aged Blue Lias belonging to the Rhomaleosauridae. Early plesiosaurs were generally small-bodied forms, with body size increasing into the Toarcian.[148] The Middle Jurassic saw the evolution of short necked and large headed thalassophonean pliosaurs from ancestrally small-headed long necked forms.[149] Some thalassophonean pliosaurs, such as some species of Pliosaurus had skulls up to two metres in length with body lengths estimated around 10–12 metres, making them among the largest predators of Jurassic oceans.[150] Cryptoclididae, a family of long-necked plesiosaurs, would radiate beginning in the Middle Jurassic.[151] Plesiosaurs would invade freshwater environments during the Jurassic, with indeterminate remains of small bodied pleisosaurs known from freshwater sediments from the Jurassic of China and Australia.[152][153]
Pterosaurs
Pterosaurs first appeared in the Late Triassic. A major radiation of Jurassic pterosaurs is the Rhamphorhynchidae, which first appeared in the late Early Jurassic (Toarcian),[154] and are thought to have had piscivorous habits.[155]Anurognathids, which first appeared in the Middle Jurassic, possess short heads and densely furred bodies, and are thought to have been insectivores.[155] Derived monofenestratan pterosaurs, like wukongopterids appeared in the late Middle Jurassic. Advanced short-tailed pterodactyloids first appeared at the Middle-Late Jurassic boundary. Jurassic pterodactyloids include the ctenochasmatids like Ctenochasma, which have closely spaced needle like teeth that were presumably used for filter feeding.[155] The bizarre Late Jurassic ctenochasmatoid Cycnorhamphus possessed a jaw with teeth only at the tips, with bent jaws like those of living openbill storks, which may have been used to hold and crush hard invertebrates.[155]
Dinosaurs
Dinosaurs, which had diversified in the Late Triassic, would experience a radiation during the Early Jurassic in the aftermath of the end Triassic extinction and the extinction of other reptile groups, becoming the dominant vertebrates in terrestrial ecosystems.[156] Chilesaurus, a morphologically aberrant herbivorous dinosaur from the Late Jurassic of South America, has uncertain relationships to the three main groups of dinosaurs, having been recovered as a member of all three in different analyses.[157]
Theropods
Advanced theropods belonging to Neotheropoda first appeared in the Late Triassic. Basal neotheropods, such as coelophysoids and "dilophosaurs", perisisted into the Early Jurassic.[158] The earliest averostrans appear during the Early Jurassic, with the earliest known member of Ceratosauria being Saltriovenator from the early Sinemurian (199.3–197.5 million years ago) of Italy.[159] The unusual ceratosaur Limusaurus from the Late Jurassic of China had a herbivorous diet with adults having edentuous beaked jaws,[160] making it the earliest known theropod to have converted from an ancestrally carnivorous diet.[161] The earliest members of the Tetanurae appeared during the early Middle Jurassic.[162] The Megalosauridae represent the oldest radiation of the Tetanurae, first appearing in Europe during the Bajocian.[163] The oldest member of Allosauroidea has been suggested to be Asfaltovenator from the Middle Jurassic of South America.[164] Coelurosaurs first appeared during the Middle Jurassic, including early tyrannosaurs such as Proceratosaurus from the Bathonian of Britain.[165] The earliest alvarezsaurs, a clade of coelurosaurs, first appeared during the Late Jurassic, represented by Shishugounykus and Haplocheirus from the Late Jurassic of China.[166] Scansoriopterygids, a group of small feathered coelurosaurs with membraneous, bat like wings for gliding, are known from the Middle-Late Jurassic of China.[167] The oldest record of troodontids is suggested to be Hesperornithoides from the Late Jurassic of North America. Tooth remains suggested to represent those of dromaeosaurs are known from the Jurassic, but no body remains are known until the Cretaceous.[168]
 Skeleton of Ceratosaurus, a ceratosaurid from the Late Jurassic of North America.
Skeleton of Ceratosaurus, a ceratosaurid from the Late Jurassic of North America. Skeleton of Monolophosaurus, a basal tetanuran from the Middle Jurassic of China
Skeleton of Monolophosaurus, a basal tetanuran from the Middle Jurassic of China Restoration of Yi qi, a scansoriopterygid from the Middle-Late Jurassic of China
Restoration of Yi qi, a scansoriopterygid from the Middle-Late Jurassic of China
Birds
.jpg.webp)
The earliest avialans, which include birds and their ancestors, appear during the Middle-Late Jurassic, definitively represented by Archaeopteryx from the Late Jurassic of Germany. Avialans belong to the clade Paraves within Coelurosuria, which also includes dromaeosaurs and troodontids. The Anchiornithidae from the Middle-Late Jurassic of China have frequently suggested to be avialans, but have also been found as a separate lineage of paravians.[162]
Ornithischians
The earliest definitive ornithischians appear during the Early Jurassic, represented by basal ornithischians like Lesothosaurus, heterodontosaurids, and early members of Thyreophora. The earliest members of Ankylosauria and Stegosauria appear during the Middle Jurassic.[169] The basal neornithischian Kulindadromeus from the Middle Jurassic of Russia indicates that at least some ornithischians were covered in protofeathers.[170] The earliest members of Ankylopollexia, which would become prominent in the Cretaceous, appeared during the Late Jurassic, represented by bipedal forms such as Camptosaurus.[171] Ceratopsians first appeared in the Late Jurassic of China, represented by members of Chaoyangsauridae.[172]
Sauropodomorphs
Bipedal basal sauropodomorphs, such as massospondylids, continued to exist into the Early Jurassic. Quadrupedal sauropomorphs appeared during the Late Triassic. The quadupedal Ledumahadi from the earliest Jurassic of South Africa reached an estimated weight of 12 tons, far in excess of other known basal sauropodomorphs.[173] Gravisaurian sauropods first appeared during the Early Jurassic, with the oldest definitive record being Vulcanodon from Zimbabwe, likely of Sinemurian age.[174] Eusauropodan sauropods diverisfied during the Middle Jurassic, including cetiosaurids, turiasaurs,[175] and mamenchisaurs.[176] The earliest known member of Neosauropoda is Lingwulong, a dicraeosaurid diplodocoid from the Middle Jurassic of China. Neosauropods dispersed worldwide during the Late Jurassic.[177] The Late Jurassic saw the first appearance of several derived neosauropod groups, including Brachiosauridae[178] and Diplodocidae.[179] Jurassic sauropods would reach gigantic sizes, becoming the largest organisms to have ever lived on land up to that point.[180]
Amphibians

The vast majority of temnospondyls went extinct at the end of the Triassic, with only brachyopoids surviving into the Jurassic and beyond. Members of the family Brachyopidae are known from Jurassic deposits in Asia,[181] while the chigutisaurid Siderops is known from the Early Jurassic of Australia.[182] Modern lissamphibians would also begin to diversify during the Jurassic. The Early Jurassic Prosalirus thought to represent the first frog relative with a morphology capable of hopping like living frogs.[183] Morphogically recognisable stem-frogs like Notobatrachus are known from the Middle Jurassic.[184] While the earliest salamander-line amphibians are known from the Triassic.[185] Crown group salamanders first appear during the Middle-Late Jurassic of Eurasia, alongside stem-group relatives. Many Jurassic stem-group salamanders, like Marmorerpeton and Kokartus are thought to have been neotenic.[186] Early representatives of crown group salamanders include Chunerpeton, Pangerpeton and Linglongtriton from the Middle-Late Jurassic Yanliao Biota of China, which belong to the Cryptobranchoidea, which contains living Asiatic and giant salamanders.[187] While Beiyanerpeton, also from the same biota is thought to be an early member of Salamandroidea, the group which contains all other living salamanders.[188] Salamanders would disperse into North America by the end of the Jurassic, as evidenced by Iridotriton found in the Late Jurassic Morrison Formation.[189] The oldest undisputed stem-caecilian is the Early Jurassic Eocaecilia from Arizona.[190] The fourth group of lissamphibians, the extinct albanerpetontids, would first appear in the Middle Jurassic, represented by Anoualerpeton priscus from the Bathonian of Britain, as well as indeterminate remains from equivalently aged sediments in France and the Anoual Formation of Morocco.[191]
Mammals
Mammals, having originated from cynodonts at the end of the Triassic, diversified extensively during the Jurassic. Important groups of Jurassic mammals include Morganucodonta, Docodonta, Eutriconodonta, Dryolestida, Haramiyida and Multituberculata. While most Jurassic mammals are solely known from isolated teeth and jaw fragments, exceptionally preserved remains have revealed a variety of lifestyles.[192] The docodontan Castorocauda was adapted for aquatic life, similar to the platypus and otters.[193] Some members of Haramiyida[194] and the eutriconodontan tribe Volaticotherini[195] possessed a patagium akin to those of flying squirrels, allowing them to glide through the air. The aardvark like mammal Fruitafossor of uncertain affinities was likely a specialist on colonial insects, similar to living anteaters.[196] Early relatives of monotremes, the Australosphenida, first appear in the Middle Jurassic of Gondwana.[197] Theriiform mammals, represented today by living placentals and marsupials, would also appear during the early Late Jurassic, represented by Juramaia, a eutherian mammal closer to the ancestry of placentals than marsupials.[198] Juramaia is much more advanced than expected for its age, as other theriiform mammals do not appear until the Early Cretaceous.[199] Non-mammalian cynodonts of the families Tritheledontidae and Tritylodontidae survived the T-J extinction, and continued to exist into the Early Jurassic and Early Cretaceous, respectively.[200]
Conodonts
The last known species of conodont, a class of jawless fish-like animals whose hard teeth like elements are key index fossils, would finally become extinct during the earliest Jurassic after over 300 million years of evolutionary history, with an asynchronous extinction occurring first in the Tethys and eastern Panthalassia, with late survivors in the earliest Hettangian of Hungary and central Panthalassa.[201] End-Triassic conodonts were only represented by a handful of species, and had been progressively declining though the Middle and Late Triassic.[202]
Sarcopterygii

Lungfish (Dipnoi) were present in freshwater environments of both hemispheres during the Jurassic. Genera include Ceratodus and Ptychoceratodus which are more closely related to living South American and African lungfish than Queensland lungfish and Ferganoceratodus from the Jurassic of Asia, which is not closely related to either group of living lungfish.[203] Mawsoniids, a marine and freshwater/brackish group of coelacanths, which first appeared in North America during the Triassic, would expand into Europe and South America by the end of the Jurassic.[204] The marine Latimeriidae, which contains the living coelacanths of the genus Latimeria, were also present in the Jurassic, having originated in the Triassic.[205]
Actinopterygii
.png.webp)
Bony fish (Actinopterygii) were major components of Jurassic freshwater and marine ecosystems. Amiiform fish (which today only includes the bowfin) would first appear during the Early Jurassic, represented by Caturus from the Pliensbachian of Britain, after their first appearance in western Tethys, they would expand to Africa, North America and Southeast and East Asia by the end of the Jurassic. Pycnodontiformes, which first appeared in the western Tethys during the Late Triassic, would expand to South America and Southeast Asia by the end of the Jurassic, having a high diversity in Europe during the Late Jurassic.[206] Teleosts, which make up over 99% of living Actinopterygii, had first appeared during the Triassic in the western Tethys, underwent a major diversification beginning in the Late Jurassic, with early representatives of modern teleost clades such as Elopomorpha and Osteoglossoidei appearing during this time.[207][208] The Pachycormiformes, a group of fish closely allied to teleosts, first appeared in the Early Jurassic, and included both tuna-like predatory and filter feeding forms, the latter including the largest bony fish known to have existed, Leedsichthys, with an estimated maximum length of over 15 metres, known from the late Middle to Late Jurassic.[209]
Chondrichthyes

The Jurassic was a major time in the evolution of Chondrichthyes. Hybodonts, such as Hybodus were common in the Early Jurassic in both marine and freshwater settings, however by the Late Jurassic, hybodonts had become minor components of most marine communities, having been largely replaced by modern neoselachians, but would remain common in freshwater and restricted marine environments.[210] The Neoselachii, which contains all living sharks and rays, would radiate beginning in the Early Jurassic.[211] The oldest known Hexanchiformes are from the Early Jurassic (Pliensbachian) of Europe.[212] The oldest known ray (Batoidea) is Antiquaobatis from the Pliensbachian of Germany.[213] Jurassic batoids known from complete remains retain a conservative, guitarfish-like morphology.[214] The oldest known relatives of the Bullhead shark (Heterodontus) in the order Heterodontiformes first appear in the Early Jurassic, with representatives of the living genus appearing during the Late Jurassic.[215] Carpet sharks (Orectolobiformes) first appear during the Toarcian, represented by Folipistrix and Annea from Europe.[216] The oldest known mackerel sharks (Lamniformes) are known from the Middle Jurassic, represented by the genus Palaeocarcharias., which has an orectolobiform-like bodyform, but shares key similarities in tooth histology with lamniformes, including the absence of orthodentine.[217] The oldest record of Angelsharks (Squatiniformes) is Pseudorhina from the Oxfordian-Tithonian of Europe.[218] The oldest known remains of Carcharhiniformes, the largest order of living sharks, first appear in the late Middle Jurassic (Bathonian) of the western Tethys (England and Morocco). Known dental and exceptionally preserved body remains of Jurassic Carchariniformes are similar to those of living catsharks.[219] The oldest remains of chimaeras are known from the Early Jurassic of Europe, with members of the living family Callorhinchidae appearing during the Middle Jurassic. Unlike living chimaeras, these were found in shallow water settings.[220] The closely related Squaloraja and myriacanthoids are also known from the Jurassic of Europe.[221]
Insects and arachnids
Numerous important insect fossil localities are known from the Jurassic of Eurasia, the most important being the Karabastau Formation of Kazakhstan, and the various Yanliao Biota deposits in Inner Mongolia, China, such as the Daohugou Bed, dating to the Callovian-Oxfordian. The diversity of insects was stagnant throughout the Early and Middle Jurassic, but during the latter third of the Jurassic origination rates increased substantially while extinction rates remained flat.[222] The Middle-Late Jurassic was a time of major diverisification for beetles.[223] Weevils first appear in the fossil record during the Middle-Upper Jurassic, but are suspected to have originated during the Late Triassic-Early Jurassic.[224] The oldest known lepidopterans (the group containing butterflies and moths) are known from the Triassic-Jurassic boundary, with wing scales belonging to the suborder Glossata and Micropterigidae-grade moths from the deposits of this age in Germany.[225] Although modern representatives are not known until the Cenozoic, ectoparasitic insects thought to represent a stem-group to fleas first appear during the Jurassic, such as Pseudopulex jurassicus. These insects are substantially different from modern fleas, lacking the specialised morphology of modern fleas and being larger in size.[226][227] The earliest group of Phasmatodea (stick insects), the winged Susumanioidea, an outgroup to living Phasmatodeans, first appear during the Middle Jurassic.[228] The oldest member of the Mantophasmatidae (gladiators) also appeared during this time.[229]
Only a handful of records of mites are known from the Jurassic, including Jureremus, an Oribatid mite belonging to the family Cymbaeremaeidae known from the Upper Jurassic of Britain and Russia.[230] Spiders would diversify through the Jurassic.[231] The Early Jurassic Seppo koponeni thought to possibly represent a stem-group to Palpimanoidea.[232] Eoplectreurys from the Middle Jurassic of China is considered a stem lineage of Synspermiata. The oldest member of the family Archaeidae, Patarchaea, is known from the Middle Jurassic.[231] Mongolarachne from the Middle Jurassic of China is among the largest known fossil spiders, with legs over 5 centimetres in length.[233] The only scorpion known from the Jurassic is Liassoscorpionides from the Lower Jurassic of Germany, of uncertain placement.[234] Eupnoi Opiliones are known from the Middle Jurassic, including members of the family Sclerosomatidae.[235][236]
End-Triassic extinction
During the end-Triassic extinction, 46%-72% of all marine genera would become extinct. The effects of the end Triassic extinction were greatest at tropical latitudes, and were more severe in Panthalassa than the Tethys or Boreal oceans. Tropical reef ecosystems would completely collapse during the event, and would not fully recover until much later in the Jurassic. Sessile filter feeders and photosymbiotic organisms were among most severely affected.[237]
Marine ecosystems
Having declined at the T-J boundary. Reefs would substantially expand during the Late Jurassic, including both sponge reefs and scleractinian coral reefs. Late Jurassic reefs were similar in form to modern reefs, but had more microbial carbonates and hypercalcified sponges, and had weak biogenic binding. Reefs would sharply decline at the close of the Jurassic.[238] which caused an associated drop in diversity in decapod crustaceans.[239] The earliest planktonic foraminifera are known from the late Early Jurassic (mid Toarcian) of the western Tethys, expanding across the whole Tethys by the Middle Jurassic, before being globally distributed in tropical latitudes by the Late Jurassic.[240] Coccolithophores and dinoflagellates, which had first appeared during the Triassic, would radiate during the Early-Middle Jurassic, becoming prominent members of the phytoplankton.[241] Microconchid tube worms, the last remaining order of Tentaculita, a group of animals of uncertain affinites that were convergent on Spirorbis tube worms, had become rare after the Triassic, and had become reduced to the single genus Punctaconchus, which became extinct in the late Bathonian.[242] The oldest known diatom is known from amber found in the Late Jurassic of Thailand, assigned to the living genus Hemiaulus.[243]
Crustaceans
The Jurassic was a significant time for the evolution of decapods.[239] The first true crabs (Brachyura) would appear during the Early Jurassic, with the earliest being Eocarcinus praecursor from the early Pliensbachian of England, which lacked the carcinisation of modern crabs.[244] and Eoprosopon klugi from the late Pliensbachian of Germany, which possibly belongs to the living family Homolodromiidae.[245] Most Jurassic crabs are only known from dorsal carapace pieces, which makes it difficult to determine their relationships.[246] While rare in the Early and Middle Jurassic, crabs would become abundant during the Late Jurassic as they expanded from their ancestral silty sea floor habitat into hard substrate habitats like reefs, with crevices in reefs helping to hide from predators[246][239] Hermit crabs would also first appear during the Jurassic, with the earliest known being Schobertella hoelderi from the late Hettangian of Germany.[247] Early hermit crabs are associated with ammonite shells rather than those of gastropods.[248] Glypheids, which today are only known from two species, reached their peak diversity during the Jurassic, with around 150 species out of a total fossil record of 250 being known from the period.[249] Thylacocephalans are an enigmatic group of arthropods known from the Silurian to the Cretaceous, with probable crustacean affinities. Remains of Jurassic thylacocephalans are among the best preserved members of the group.[250]
Brachiopods
Brachiopod diversity declined during the T-J extinction. Spire bearing groups (Spiriferinida and Athyridida) would significantly decline at the T-J boundary and would not recover their biodiversity, becoming extinct in the TOAE.[48] Rhynchonellida and Terebratulida would also decline during the T-J extincton but would rebound during the Early Jurassic, neither clade would develop any significant morphological variation.[251] Brachiopods would substantially decline in the Late Jurassic, the causes of which are poorly understood. Proposed reasons include increased predation, competition with bivalves, enhanced bioturbation or increased grazing pressure.[252]
Ammonites
Ammonites were devastated by the end-Triassic extinction, with only the family Psiloceratidae of the suborder Phylloceratina surviving, and becoming ancestral to all later Jurassic and Cretaceous ammonites. Ammonites explosively diversified during the Early Jurassic, with the orders Psiloceratina, Ammonitina, Lytoceratina, Haploceratina, Perisphinctina and Ancyloceratina all appearing during the Jurassic. Ammonite faunas during the Jurassic were regional, being divided into around 20 distinguishable provinces and subprovinces in two realms, the northern high latitude Pan-Boreal realm, consisting of the Arctic, northern Panthalassa and northern Atlantic regions, and the equatorial-southern Pan-Tethyan realm, which included the Tethys and most of Panthalassa.[253]
Bivalves
The end Triassic extinction had a severe impact on bivalve diversity, though it had little impact on bivalve ecological diversity. The extinction was selective, having less of an impact on deep burrowers, but there is no evidence of a differential impact between surface living (epifaunal) and burrowing (infaunal) bivalves.[254] Rudists, the dominant reef building organisms of the Cretaceous, would first appear in the Late Jurassic (mid Oxfordian) in the northern margin of the western Tethys region, expanding to the eastern Tethys by the end of the Jurassic.[255]
Belemnites
The oldest definitive records of the squid-like belemnites are from the earliest Jurassic (Hettangian-Sinemurian) of Europe and Japan, and would expand worldwide during the Jurassic.[256] Belemnites were shallow water dwellers, inhabiting the upper 200 metres of the water column on the continental shelves and in the littoral zone. They were key components of Jurassic ecosystems, both as predators and prey, as evidenced by the abundance of belemnite guards in Jurassic rocks.[257]
References
Notes
- A 140 Ma age for the Jurassic-Cretaceous instead of the usually accepted 145 Ma was proposed in 2014 based on a stratigraphic study of Vaca Muerta Formation in Neuquén Basin, Argentina.[2] Víctor Ramos, one of the authors of the study proposing the 140 Ma boundary age, sees the study as a "first step" toward formally changing the age in the International Union of Geological Sciences.[3]
- "Ich hatte mich auf einer geognostischen Reise, die ich 1795 durch das südliche Franken, die westliche Schweiz und Ober-Italien machte, davon überzeugt, daß der Jura-Kalkstein, welchen Werner zu seinem Muschelkalk rechnete, eine eigne Formation bildete. In meiner Schrift über die unterirdischen Gasarten, welche mein Bruder Wilhelm von Humboldt 1799 während meines Aufenthalts in Südamerika herausgab, wird der Formation, die ich vorläufig mit dem Namen Jura-Kalkstein bezeichnete, zuerst gedacht." ('On a geological tour that I made in 1795 through southern France, western Switzerland and upper Italy, I convinced myself that the Jura limestone, which Werner included in his shell limestone, constituted a separate formation. In my paper about subterranean types of gases, which my brother Wilhelm von Humboldt published in 1799 during my stay in South America, the formation, which I provisionally designated with the name "Jura limestone", is first conceived.')[8]
- "[…] die ausgebreitete Formation, welche zwischen dem alten Gips und neueren Sandstein liegt, und welchen ich vorläufig mit dem Nahmen Jura-Kalkstein bezeichne." '… the widespread formation which lies between the old gypsum and the more recent sandstone and which I provisionally designate with the name "Jura limestone".'[9]
Citations
- "Jurassic". Lexico Dictionaries. Retrieved 2021-01-09.
- Vennari et al. 2014, pp. 374-385.
- Jaramillo 2014.
- Hallam 1986, pp. 765-768.
- Hölder 1964.
- Arkell 1956.
- Rollier 1903.
- von Humboldt 1858, p. 632.
- von Humboldt 1799, p. 39.
- Pieńkowski et al. 2008, pp. 823–922.
- Brongniart 1829.
- von Buch, L., 1839. Über den Jura in Deutschland. Der Königlich Preussischen Akademie der Wissenschaften, Berlin, p. 87.
- Ogg, J.G.; Hinnov, L.A.; Huang, C. (2012), "Jurassic", The Geologic Time Scale, Elsevier, pp. 731–791, doi:10.1016/b978-0-444-59425-9.00026-3, ISBN 978-0-444-59425-9, retrieved 2020-12-05
- Kazlev 2002.
- Hillebrandt, A.v.; Krystyn, L.; Kürschner, W.M.; Bonis, N.R.; Ruhl, M.; Richoz, S.; Schobben, M. A. N.; Urlichs, M.; Bown, P.R.; Kment, K.; McRoberts, C.A. (2013-09-01). "The Global Stratotype Sections and Point (GSSP) for the base of the Jurassic System at Kuhjoch (Karwendel Mountains, Northern Calcareous Alps, Tyrol, Austria)". Episodes. 36 (3): 162–198. doi:10.18814/epiiugs/2013/v36i3/001. ISSN 0705-3797.
- Bloos, Gert; Page, Kevin N. (2002-03-01). "Global Stratotype Section and Point for base of the Sinemurian Stage (Lower Jurassic)". Episodes. 25 (1): 22–28. doi:10.18814/epiiugs/2002/v25i1/003. ISSN 0705-3797.
- Meister, Christian; Aberhan, Martin; Blau, Joachim; Dommergues, Jean-Louis; Feist-Burkhardt, Susanne; Hailwood, Ernie A.; Hart, Malcom; Hesselbo, Stephen P.; Hounslow, Mark W.; Hylton, Mark; Morton, Nicol (2006-06-01). "The Global Boundary Stratotype Section and Point (GSSP) for the base of the Pliensbachian Stage (Lower Jurassic), Wine Haven, Yorkshire, UK". Episodes. 29 (2): 93–106. doi:10.18814/epiiugs/2006/v29i2/003. ISSN 0705-3797.
- Fantasia, Alicia; Adatte, Thierry; Spangenberg, Jorge E.; Font, Eric; Duarte, Luís V.; Föllmi, Karl B. (November 2019). "Global versus local processes during the Pliensbachian–Toarcian transition at the Peniche GSSP, Portugal: A multi-proxy record". Earth-Science Reviews. 198: 102932. Bibcode:2019ESRv..19802932F. doi:10.1016/j.earscirev.2019.102932.
- Barrón, Eduardo; Ureta, Soledad; Goy, Antonio; Lassaletta, Luis (August 2010). "Palynology of the Toarcian–Aalenian Global Boundary Stratotype Section and Point (GSSP) at Fuentelsaz (Lower–Middle Jurassic, Iberian Range, Spain)". Review of Palaeobotany and Palynology. 162 (1): 11–28. doi:10.1016/j.revpalbo.2010.04.003.
- Pavia, G.; Enay, R. (1997-03-01). "Definition of the Aalenian-Bajocian Stage boundary". Episodes. 20 (1): 16–22. doi:10.18814/epiiugs/1997/v20i1/004. ISSN 0705-3797.
- López, Fernández; Rafael, Sixto; Pavia, Giulio; Erba, Elisabetta; Guiomar, Myette; Paiva Henriques, María Helena; Lanza, Roberto; Mangold, Charles; Morton, Nicol; Olivero, Davide; Tiraboschi, Daniele (2009). "The Global Boundary Stratotype Section and Point (GSSP) for base of the Bathonian Stage (Middle Jurassic), Ravin du Bès Section, SE France" (PDF). Episodes. 32 (4): 222–248. doi:10.18814/epiiugs/2009/v32i4/001. Archived from the original (PDF) on 4 March 2016. Retrieved 5 June 2015.
- BARSKI, Marcin (2018-09-06). "Dinoflagellate cyst assemblages across the Oxfordian/Kimmeridgian boundary (Upper Jurassic) at Flodigarry, Staffin Bay, Isle of Skye, Scotland – a proposed GSSP for the base of the Kimmeridgian". Volumina Jurassica. XV (1): 51–62. doi:10.5604/01.3001.0012.4594. ISSN 1731-3708.
- WIMBLEDON, William A.P. (2017-12-27). "Developments with fixing a Tithonian/Berriasian (J/K) boundary". Volumina Jurassica (1): 0. doi:10.5604/01.3001.0010.7467. ISSN 1731-3708.
- Gautier D.L. (2005). "Kimmeridgian Shales Total Petroleum System of the North Sea Graben Province" (PDF). United States Geological Survey. Retrieved 2 November 2018.
- Wilson, A. O. (2020). "Chapter 1 Introduction to the Jurassic Arabian Intrashelf Basin". Geological Society, London, Memoirs. 53 (1): 1–19. doi:10.1144/M53.1. ISSN 0435-4052. S2CID 226967035.
- Abdula, Rzger A. (August 2015). "Hydrocarbon potential of Sargelu Formation and oil-source correlation, Iraqi Kurdistan". Arabian Journal of Geosciences. 8 (8): 5845–5868. doi:10.1007/s12517-014-1651-0. ISSN 1866-7511. S2CID 129120960.
- Soran University; Abdula, Rzger A. (2016-10-16). "Source Rock Assessment of Naokelekan Formation in Iraqi Kurdistan". Journal of Zankoy Sulaimani - Part A. 19 (1): 103–124. doi:10.17656/jzs.10589.
- Ao, Weihua; Huang, Wenhui; Weng, Chengmin; Xiao, Xiuling; Liu, Dameng; Tang, Xiuyi; Chen, Ping; Zhao, Zhigen; Wan, Huan; Finkelman, Robert B. (January 2012). "Coal petrology and genesis of Jurassic coal in the Ordos Basin, China". Geoscience Frontiers. 3 (1): 85–95. doi:10.1016/j.gsf.2011.09.004.
- Tennant, Jonathan P.; Mannion, Philip D.; Upchurch, Paul (2016-09-02). "Sea level regulated tetrapod diversity dynamics through the Jurassic/Cretaceous interval". Nature Communications. 7 (1): 12737. Bibcode:2016NatCo...712737T. doi:10.1038/ncomms12737. ISSN 2041-1723. PMC 5025807. PMID 27587285.
- Holm‐Alwmark, S.; Alwmark, C.; Ferrière, L.; Lindström, S.; Meier, M. M. M.; Scherstén, A.; Herrmann, M.; Masaitis, V. L.; Mashchak, M. S.; Naumov, M. V.; Jourdan, F. (August 2019). "An Early Jurassic age for the Puchezh‐Katunki impact structure (Russia) based on 40 Ar/ 39 Ar data and palynology". Meteoritics & Planetary Science. 54 (8): 1764–1780. Bibcode:2019M&PS...54.1764H. doi:10.1111/maps.13309. ISSN 1086-9379.
- Scotese 2003.
- Barth, G.; Franz, M.; Heunisch, C.; Ernst, W.; Zimmermann, J.; Wolfgramm, M. (2018-01-01). "Marine and terrestrial sedimentation across the T–J transition in the North German Basin". Palaeogeography, Palaeoclimatology, Palaeoecology. 489: 74–94. Bibcode:2018PPP...489...74B. doi:10.1016/j.palaeo.2017.09.029. ISSN 0031-0182.
- Korte, Christoph; Hesselbo, Stephen P.; Ullmann, Clemens V.; Dietl, Gerd; Ruhl, Micha; Schweigert, Günter; Thibault, Nicolas (December 2015). "Jurassic climate mode governed by ocean gateway". Nature Communications. 6 (1): 10015. Bibcode:2015NatCo...610015K. doi:10.1038/ncomms10015. ISSN 2041-1723. PMC 4682040. PMID 26658694.
- Geiger, Markus; Clark, David Norman; Mette, Wolfgang (March 2004). "Reappraisal of the timing of the breakup of Gondwana based on sedimentological and seismic evidence from the Morondava Basin, Madagascar". Journal of African Earth Sciences. 38 (4): 363–381. Bibcode:2004JAfES..38..363G. doi:10.1016/j.jafrearsci.2004.02.003.
- Nguyen, Luan C.; Hall, Stuart A.; Bird, Dale E.; Ball, Philip J. (June 2016). "Reconstruction of the East Africa and Antarctica continental margins: AFRICA-ANTARCTICA RECONSTRUCTION". Journal of Geophysical Research: Solid Earth. 121 (6): 4156–4179. doi:10.1002/2015JB012776.
- Iturralde-Vinent, Manuel A. (2003-01-01). "The Conflicting Paleontologic versus Stratigraphic Record of the Formation of the Caribbean Seaway". The Circum-Gulf of Mexico and the Caribbean: Hydrocarbon Habitats, Basin Formation and Plate Tectonics.
- Boschman, Lydian M.; van Hinsbergen, Douwe J. J. (July 2016). "On the enigmatic birth of the Pacific Plate within the Panthalassa Ocean". Science Advances. 2 (7): e1600022. Bibcode:2016SciA....2E0022B. doi:10.1126/sciadv.1600022. ISSN 2375-2548. PMC 5919776. PMID 29713683.
- Danise, Silvia; Holland, Steven M. (July 2018). "A Sequence Stratigraphic Framework for the Middle to Late Jurassic of the Sundance Seaway, Wyoming: Implications for Correlation, Basin Evolution, and Climate Change". The Journal of Geology. 126 (4): 371–405. Bibcode:2018JG....126..371D. doi:10.1086/697692. ISSN 0022-1376. S2CID 133707199.
- Haq, Bilal U. (2018-01-01). "Jurassic Sea-Level Variations: A Reappraisal". GSA Today: 4–10. doi:10.1130/GSATG359A.1.
- Stanley & Hardie 1998.
- Eichenseer, Kilian; Balthasar, Uwe; Smart, Christopher W.; Stander, Julian; Haaga, Kristian A.; Kiessling, Wolfgang (August 2019). "Jurassic shift from abiotic to biotic control on marine ecological success". Nature Geoscience. 12 (8): 638–642. doi:10.1038/s41561-019-0392-9. hdl:10026.1/14472. ISSN 1752-0894. S2CID 197402218.
- Monroe & Wicander 1997, p. 607.
- Jacobs 1997, pp. 2-4.
- Sellwood, Bruce W.; Valdes, Paul J. (2008). "Jurassic climates". Proceedings of the Geologists' Association. 119 (1): 5–17. doi:10.1016/S0016-7878(59)80068-7.
- Them, T.R.; Gill, B.C.; Caruthers, A.H.; Gröcke, D.R.; Tulsky, E.T.; Martindale, R.C.; Poulton, T.P.; Smith, P.L. (February 2017). "High-resolution carbon isotope records of the Toarcian Oceanic Anoxic Event (Early Jurassic) from North America and implications for the global drivers of the Toarcian carbon cycle". Earth and Planetary Science Letters. 459: 118–126. Bibcode:2017E&PSL.459..118T. doi:10.1016/j.epsl.2016.11.021.
- Dera, Guillaume; Neige, Pascal; Dommergues, Jean-Louis; Fara, Emmanuel; Laffont, Rémi; Pellenard, Pierre (January 2010). "High-resolution dynamics of Early Jurassic marine extinctions: the case of Pliensbachian–Toarcian ammonites (Cephalopoda)". Journal of the Geological Society. 167 (1): 21–33. Bibcode:2010JGSoc.167...21D. doi:10.1144/0016-76492009-068. ISSN 0016-7649. S2CID 128908746.
- Caruthers, Andrew H.; Smith, Paul L.; Gröcke, Darren R. (September 2013). "The Pliensbachian–Toarcian (Early Jurassic) extinction, a global multi-phased event". Palaeogeography, Palaeoclimatology, Palaeoecology. 386: 104–118. Bibcode:2013PPP...386..104C. doi:10.1016/j.palaeo.2013.05.010.
- Vörös, Attila; Kocsis, Ádám T.; Pálfy, József (September 2016). "Demise of the last two spire-bearing brachiopod orders (Spiriferinida and Athyridida) at the Toarcian (Early Jurassic) extinction event". Palaeogeography, Palaeoclimatology, Palaeoecology. 457: 233–241. Bibcode:2016PPP...457..233V. doi:10.1016/j.palaeo.2016.06.022.
- Maxwell, Erin E.; Vincent, Peggy (2015-11-06). "Effects of the early Toarcian Oceanic Anoxic Event on ichthyosaur body size and faunal composition in the Southwest German Basin". Paleobiology. 42 (1): 117–126. doi:10.1017/pab.2015.34. ISSN 0094-8373. S2CID 131623205.
- Xu, Weimu; Ruhl, Micha; Jenkyns, Hugh C.; Hesselbo, Stephen P.; Riding, James B.; Selby, David; Naafs, B. David A.; Weijers, Johan W. H.; Pancost, Richard D.; Tegelaar, Erik W.; Idiz, Erdem F. (February 2017). "Carbon sequestration in an expanded lake system during the Toarcian oceanic anoxic event". Nature Geoscience. 10 (2): 129–134. Bibcode:2017NatGe..10..129X. doi:10.1038/ngeo2871. ISSN 1752-0894.
- Müller, Tamás; Jurikova, Hana; Gutjahr, Marcus; Tomašových, Adam; Schlögl, Jan; Liebetrau, Volker; Duarte, Luís v.; Milovský, Rastislav; Suan, Guillaume; Mattioli, Emanuela; Pittet, Bernard (2020-12-01). "Ocean acidification during the early Toarcian extinction event: Evidence from boron isotopes in brachiopods". Geology. 48 (12): 1184–1188. Bibcode:2020Geo....48.1184M. doi:10.1130/G47781.1. ISSN 0091-7613.
- Trecalli, Alberto; Spangenberg, Jorge; Adatte, Thierry; Föllmi, Karl B.; Parente, Mariano (December 2012). "Carbonate platform evidence of ocean acidification at the onset of the early Toarcian oceanic anoxic event". Earth and Planetary Science Letters. 357–358: 214–225. Bibcode:2012E&PSL.357..214T. doi:10.1016/j.epsl.2012.09.043.
- Ettinger, Nicholas P.; Larson, Toti E.; Kerans, Charles; Thibodeau, Alyson M.; Hattori, Kelly E.; Kacur, Sean M.; Martindale, Rowan C. (2020-09-23). Eberli, Gregor (ed.). "Ocean acidification and photic‐zone anoxia at the Toarcian Oceanic Anoxic Event: Insights from the Adriatic Carbonate Platform". Sedimentology. 68: 63–107. doi:10.1111/sed.12786. ISSN 0037-0746.
- Elgorriaga, Andrés; Escapa, Ignacio H.; Cúneo, N. Rubén (July 2019). "Relictual Lepidopteris (Peltaspermales) from the Early Jurassic Cañadón Asfalto Formation, Patagonia, Argentina". International Journal of Plant Sciences. 180 (6): 578–596. doi:10.1086/703461. ISSN 1058-5893. S2CID 195435840.
- Mander, Luke; Kürschner, Wolfram M.; McElwain, Jennifer C. (2010-08-31). "An explanation for conflicting records of Triassic–Jurassic plant diversity". Proceedings of the National Academy of Sciences. 107 (35): 15351–15356. Bibcode:2010PNAS..10715351M. doi:10.1073/pnas.1004207107. ISSN 0027-8424. PMC 2932585. PMID 20713737.
- Barbacka, Maria; Pacyna, Grzegorz; Kocsis, Ádam T.; Jarzynka, Agata; Ziaja, Jadwiga; Bodor, Emese (August 2017). "Changes in terrestrial floras at the Triassic-Jurassic Boundary in Europe". Palaeogeography, Palaeoclimatology, Palaeoecology. 480: 80–93. Bibcode:2017PPP...480...80B. doi:10.1016/j.palaeo.2017.05.024.
- Bomfleur, Benjamin; Blomenkemper, Patrick; Kerp, Hans; McLoughlin, Stephen (2018), "Polar Regions of the Mesozoic–Paleogene Greenhouse World as Refugia for Relict Plant Groups", Transformative Paleobotany, Elsevier, pp. 593–611, doi:10.1016/b978-0-12-813012-4.00024-3, ISBN 978-0-12-813012-4, retrieved 2020-11-12
- Bateman, Richard M (2020-01-01). Ort, Donald (ed.). "Hunting the Snark: the flawed search for mythical Jurassic angiosperms". Journal of Experimental Botany. 71 (1): 22–35. doi:10.1093/jxb/erz411. ISSN 0022-0957. PMID 31538196.
- Stockey, Ruth A.; Rothwell, Gar W. (July 2020). "Diversification of crown group Araucaria : the role of Araucaria famii sp. nov. in the Late Cretaceous (Campanian) radiation of Araucariaceae in the Northern Hemisphere". American Journal of Botany. 107 (7): 1072–1093. doi:10.1002/ajb2.1505. ISSN 0002-9122. PMID 32705687.
- Escapa, Ignacio H.; Catalano, Santiago A. (October 2013). "Phylogenetic Analysis of Araucariaceae: Integrating Molecules, Morphology, and Fossils". International Journal of Plant Sciences. 174 (8): 1153–1170. doi:10.1086/672369. hdl:11336/3583. ISSN 1058-5893. S2CID 56238574.
- Stockey, Ruth A.; Rothwell, Gar W. (March 2013). "Pararaucaria carrii sp. nov., Anatomically Preserved Evidence for the Conifer Family Cheirolepidiaceae in the Northern Hemisphere". International Journal of Plant Sciences. 174 (3): 445–457. doi:10.1086/668614. ISSN 1058-5893. S2CID 59269291.
- Escapa, Ignacio; Cúneo, Rubén; Axsmith, Brian (September 2008). "A new genus of the Cupressaceae (sensu lato) from the Jurassic of Patagonia: Implications for conifer megasporangiate cone homologies". Review of Palaeobotany and Palynology. 151 (3–4): 110–122. doi:10.1016/j.revpalbo.2008.03.002.
- Contreras, Dori L.; Escapa, Ignacio H.; Iribarren, Rocio C.; Cúneo, N. Rubén (October 2019). "Reconstructing the Early Evolution of the Cupressaceae: A Whole-Plant Description of a New Austrohamia Species from the Cañadón Asfalto Formation (Early Jurassic), Argentina". International Journal of Plant Sciences. 180 (8): 834–868. doi:10.1086/704831. ISSN 1058-5893. S2CID 202862782.
- Spencer, A. R. T.; Mapes, G.; Bateman, R. M.; Hilton, J.; Rothwell, G. W. (2015-06-01). "Middle Jurassic evidence for the origin of Cupressaceae: A paleobotanical context for the roles of regulatory genetics and development in the evolution of conifer seed cones". American Journal of Botany. 102 (6): 942–961. doi:10.3732/ajb.1500121. ISSN 0002-9122. PMID 26101419.
- Rothwell, Gar W.; Mapes, Gene; Stockey, Ruth A.; Hilton, Jason (April 2012). "The seed cone Eathiestrobus gen. nov.: Fossil evidence for a Jurassic origin of Pinaceae". American Journal of Botany. 99 (4): 708–720. doi:10.3732/ajb.1100595. PMID 22491001.
- Pole, Mike; Wang, Yongdong; Bugdaeva, Eugenia V.; Dong, Chong; Tian, Ning; Li, Liqin; Zhou, Ning (2016-12-15). "The rise and demise of Podozamites in east Asia—An extinct conifer life style". Palaeogeography, Palaeoclimatology, Palaeoecology. Mesozoic ecosystems - Climate and Biota. 464: 97–109. Bibcode:2016PPP...464...97P. doi:10.1016/j.palaeo.2016.02.037. ISSN 0031-0182.
- Dong, Chong; Shi, Gongle; Herrera, Fabiany; Wang, Yongdong; Herendeen, Patrick S; Crane, Peter R (2020-06-18). "Middle–Late Jurassic fossils from northeastern China reveal morphological stasis in the catkin-yew". National Science Review. 7 (11): 1765–1767. doi:10.1093/nsr/nwaa138. ISSN 2095-5138.
- Nosova, N. V.; Kiritchkova, A. I. (October 2008). "A new species and a new combination of the Mesozoic genus Podocarpophyllum Gomolitzky (Coniferales)". Paleontological Journal. 42 (6): 665–674. doi:10.1134/S0031030108060129. ISSN 0031-0301. S2CID 84568477.
- Harris, T.M., 1979. The Yorkshire Jurassic flora, V. Coniferales. Trustees of the British 417 Museum (Natural History), London, 166 pp.
- Reymanówna, Maria (January 1987). "A Jurassic podocarp from Poland". Review of Palaeobotany and Palynology. 51 (1–3): 133–143. doi:10.1016/0034-6667(87)90026-1.
- Zhou, Zhi-Yan (March 2009). "An overview of fossil Ginkgoales". Palaeoworld. 18 (1): 1–22. doi:10.1016/j.palwor.2009.01.001.
In the extant species G. biloba, leaves from the same tree on different shoots or from shoots at different developmental stages may be referrable to the different leaf types (morphogenera) established for fossil ginkgoaleans.
- Nosova, Natalya (October 2013). "Revision of the genus Grenana Samylina from the Middle Jurassic of Angren, Uzbekistan". Review of Palaeobotany and Palynology. 197: 226–252. doi:10.1016/j.revpalbo.2013.06.005.
- Herrera, Fabiany; Shi, Gongle; Ichinnorov, Niiden; Takahashi, Masamichi; Bugdaeva, Eugenia V.; Herendeen, Patrick S.; Crane, Peter R. (2017-03-21). "The presumed ginkgophyte Umaltolepis has seed-bearing structures resembling those of Peltaspermales and Umkomasiales". Proceedings of the National Academy of Sciences. 114 (12): E2385–E2391. doi:10.1073/pnas.1621409114. ISSN 0027-8424. PMC 5373332. PMID 28265050.
- Barbacka, Maria; Kustatscher, Evelyn; Bodor, Emese R. (2019-03-01). "Ferns of the Lower Jurassic from the Mecsek Mountains (Hungary): taxonomy and palaeoecology". PalZ. 93 (1): 151–185. doi:10.1007/s12542-018-0430-8. ISSN 1867-6812. S2CID 135415397.
- Regalado, Ledis; Schmidt, Alexander R.; Müller, Patrick; Niedermeier, Lisa; Krings, Michael; Schneider, Harald (July 2019). "Heinrichsia cheilanthoides gen. et sp. nov., a fossil fern in the family Pteridaceae (Polypodiales) from the Cretaceous amber forests of Myanmar". Journal of Systematics and Evolution. 57 (4): 329–338. doi:10.1111/jse.12514. ISSN 1674-4918. S2CID 182754946.
- Li, Chunxiang; Miao, Xinyuan; Zhang, Li-Bing; Ma, Junye; Hao, Jiasheng (January 2020). "Re-evaluation of the systematic position of the Jurassic–Early Cretaceous fern genus Coniopteris". Cretaceous Research. 105: 104136. doi:10.1016/j.cretres.2019.04.007.
- Bomfleur, Benjamin; Grimm, Guido W.; McLoughlin, Stephen (2015-06-30). "Osmunda pulchella sp. nov. from the Jurassic of Sweden—reconciling molecular and fossil evidence in the phylogeny of modern royal ferns (Osmundaceae)". BMC Evolutionary Biology. 15 (1): 126. doi:10.1186/s12862-015-0400-7. ISSN 1471-2148. PMC 4487210. PMID 26123220.
- McLoughlin, Stephen; Bomfleur, Benjamin (December 2016). "Biotic interactions in an exceptionally well preserved osmundaceous fern rhizome from the Early Jurassic of Sweden". Palaeogeography, Palaeoclimatology, Palaeoecology. 464: 86–96. Bibcode:2016PPP...464...86M. doi:10.1016/j.palaeo.2016.01.044.
- Elgorriaga, Andrés; Escapa, Ignacio H.; Bomfleur, Benjamin; Cúneo, Rubén; Ottone, Eduardo G. (February 2015). "Reconstruction and Phylogenetic Significance of a New Equisetum Linnaeus Species from the Lower Jurassic of Cerro Bayo (Chubut Province, Argentina)". Ameghiniana. 52 (1): 135–152. doi:10.5710/AMGH.15.09.2014.2758. ISSN 0002-7014. S2CID 6134534.
- Gould, R. E. 1968. Morphology of Equisetum laterale Phillips, 1829, and E. bryanii sp. nov. from the Mesozoic of south‐eastern Queensland. Australian Journal of Botany 16: 153–176.
- Elgorriaga, Andrés; Escapa, Ignacio H.; Rothwell, Gar W.; Tomescu, Alexandru M. F.; Rubén Cúneo, N. (August 2018). "Origin of Equisetum : Evolution of horsetails (Equisetales) within the major euphyllophyte clade Sphenopsida". American Journal of Botany. 105 (8): 1286–1303. doi:10.1002/ajb2.1125. PMID 30025163.
- Channing, Alan; Zamuner, Alba; Edwards, Dianne; Guido, Diego (2011). "Equisetum thermale sp. nov. (Equisetales) from the Jurassic San Agustín hot spring deposit, Patagonia: Anatomy, paleoecology, and inferred paleoecophysiology". American Journal of Botany. 98 (4): 680–697. doi:10.3732/ajb.1000211. ISSN 1537-2197. PMID 21613167.
- Lantz, Trevor C; Rothwell, Gar W; Stockey, Ruth A (September 1999). "Conantiopteris schuchmanii, gen. et sp. nov., and the Role of Fossils in Resolving the Phylogeny of Cyatheaceae s.l." Journal of Plant Research. 112 (3): 361–381. doi:10.1007/PL00013890. ISSN 0918-9440. S2CID 23368605.
- Axsmith, Brian J.; Krings, Michael; Taylor, Thomas N. (September 2001). "A filmy fern from the Upper Triassic of North Carolina (USA)". American Journal of Botany. 88 (9): 1558–1567. doi:10.2307/3558399. ISSN 0002-9122. JSTOR 3558399. PMID 21669688.
- Taylor, T (2009), "Cycadophytes", Biology and Evolution of Fossil Plants, Elsevier, pp. 703–741, doi:10.1016/b978-0-12-373972-8.00017-6, ISBN 978-0-12-373972-8, retrieved 2020-12-12
- Pott, Christian; McLoughlin, Stephen (2014-06-01). "Divaricate growth habit in Williamsoniaceae (Bennettitales): unravelling the ecology of a key Mesozoic plant group". Palaeobiodiversity and Palaeoenvironments. 94 (2): 307–325. doi:10.1007/s12549-014-0157-9. ISSN 1867-1608. S2CID 84440045.
- Labandeira, Conrad C.; Yang, Qiang; Santiago-Blay, Jorge A.; Hotton, Carol L.; Monteiro, Antónia; Wang, Yong-Jie; Goreva, Yulia; Shih, ChungKun; Siljeström, Sandra; Rose, Tim R.; Dilcher, David L. (2016-02-10). "The evolutionary convergence of mid-Mesozoic lacewings and Cenozoic butterflies". Proceedings of the Royal Society B: Biological Sciences. 283 (1824): 20152893. doi:10.1098/rspb.2015.2893. ISSN 0962-8452. PMC 4760178. PMID 26842570.
- Khramov, Alexander V.; Lukashevich, Elena D. (July 2019). "A Jurassic dipteran pollinator with an extremely long proboscis". Gondwana Research. 71: 210–215. Bibcode:2019GondR..71..210K. doi:10.1016/j.gr.2019.02.004.
- Nagalingum, N. S.; Marshall, C. R.; Quental, T. B.; Rai, H. S.; Little, D. P.; Mathews, S. (2011-11-11). "Recent Synchronous Radiation of a Living Fossil". Science. 334 (6057): 796–799. Bibcode:2011Sci...334..796N. doi:10.1126/science.1209926. ISSN 0036-8075. PMID 22021670. S2CID 206535984.
- Condamine, Fabien L; Nagalingum, Nathalie S; Marshall, Charles R; Morlon, Hélène (December 2015). "Origin and diversification of living cycads: a cautionary tale on the impact of the branching process prior in Bayesian molecular dating". BMC Evolutionary Biology. 15 (1): 65. doi:10.1186/s12862-015-0347-8. ISSN 1471-2148. PMC 4449600. PMID 25884423.
- Coiro, Mario; Pott, Christian (December 2017). "Eobowenia gen. nov. from the Early Cretaceous of Patagonia: indication for an early divergence of Bowenia?". BMC Evolutionary Biology. 17 (1): 97. doi:10.1186/s12862-017-0943-x. ISSN 1471-2148. PMC 5383990. PMID 28388891.
- Vajda, Vivi; Pucetaite, Milda; McLoughlin, Stephen; Engdahl, Anders; Heimdal, Jimmy; Uvdal, Per (August 2017). "Molecular signatures of fossil leaves provide unexpected new evidence for extinct plant relationships". Nature Ecology & Evolution. 1 (8): 1093–1099. doi:10.1038/s41559-017-0224-5. ISSN 2397-334X. PMID 29046567. S2CID 3604369.
- Spencer, Alan R. T.; Garwood, Russell J.; Rees, Andrew R.; Raine, Robert J.; Rothwell, Gar W.; Hollingworth, Neville T. J.; Hilton, Jason (2017-08-28). "New insights into Mesozoic cycad evolution: an exploration of anatomically preserved Cycadaceae seeds from the Jurassic Oxford Clay biota". PeerJ. 5: e3723. doi:10.7717/peerj.3723. ISSN 2167-8359. PMC 5578371. PMID 28875075.
- Salgado, Leonardo; Canudo, José I.; Garrido, Alberto C.; Moreno-Azanza, Miguel; Martínez, Leandro C. A.; Coria, Rodolfo A.; Gasca, José M. (2017-02-16). "A new primitive Neornithischian dinosaur from the Jurassic of Patagonia with gut contents". Scientific Reports. 7 (1): 42778. Bibcode:2017NatSR...742778S. doi:10.1038/srep42778. ISSN 2045-2322. PMC 5311864. PMID 28202910.
- Yang, Yong; Xie, Lei; Ferguson, David K. (October 2017). "Protognetaceae: A new gnetoid macrofossil family from the Jurassic of northeastern China". Perspectives in Plant Ecology, Evolution and Systematics. 28: 67–77. doi:10.1016/j.ppees.2017.08.001.
- Elgorriaga, Andrés; Escapa, Ignacio H.; Cúneo, N. Rubén (2019-09-02). "Southern Hemisphere Caytoniales: vegetative and reproductive remains from the Lonco Trapial Formation (Lower Jurassic), Patagonia". Journal of Systematic Palaeontology. 17 (17): 1477–1495. doi:10.1080/14772019.2018.1535456. ISSN 1477-2019. S2CID 92287804.
- Taylor, Edith L.; Taylor, Thomas N.; Kerp, Hans; Hermsen, Elizabeth J. (January 2006). "Mesozoic seed ferns: Old paradigms, new discoveries 1". The Journal of the Torrey Botanical Society. 133 (1): 62–82. doi:10.3159/1095-5674(2006)133[62:MSFOPN]2.0.CO;2. ISSN 1095-5674.
- Kirchner M, Müller A. 1992. Umkomasia franconica n. sp. und Pteruchus septentrionalis n. sp. Fruktifikationen von Thinnfeldia Ettinghausen. Palaeontographica 224B: 63–73.
- Kustatscher, Evelyn; Visscher, Henk; van Konijnenburg-van Cittert, Johanna H. A. (2019-09-01). "Did the Czekanowskiales already exist in the late Permian?". PalZ. 93 (3): 465–477. doi:10.1007/s12542-019-00468-9. ISSN 1867-6812. S2CID 199473893.
- Taylor, T (2009), "Gymnosperms with obscure affinities", Biology and Evolution of Fossil Plants, Elsevier, pp. 757–785, doi:10.1016/b978-0-12-373972-8.00019-x, ISBN 978-0-12-373972-8, retrieved 2020-12-13
- Wood, Daniel; Besnard, Guillaume; Beerling, David J.; Osborne, Colin P.; Christin, Pascal-Antoine (2020-06-18). "Phylogenomics indicates the "living fossil" Isoetes diversified in the Cenozoic". PLOS ONE. 15 (6): e0227525. Bibcode:2020PLoSO..1527525W. doi:10.1371/journal.pone.0227525. ISSN 1932-6203. PMC 7302493. PMID 32555586.
- Mamontov, Yuriy S.; Ignatov, Michael S. (July 2019). "How to rely on the unreliable: Examples from Mesozoic bryophytes of Transbaikalia". Journal of Systematics and Evolution. 57 (4): 339–360. doi:10.1111/jse.12483. ISSN 1674-4918. S2CID 92268163.
- Bippus, Alexander C.; Savoretti, Adolfina; Escapa, Ignacio H.; Garcia-Massini, Juan; Guido, Diego (October 2019). "Heinrichsiella patagonica gen. et sp. nov.: A Permineralized Acrocarpous Moss from the Jurassic of Patagonia". International Journal of Plant Sciences. 180 (8): 882–891. doi:10.1086/704832. ISSN 1058-5893. S2CID 202859471.
- Li, Ruiyun; Li, Xiaoqiang; Deng, Shenghui; Sun, Bainian (August 2020). "Morphology and microstructure of Pellites hamiensis nov. sp., a Middle Jurassic liverwort from northwestern China and its evolutionary significance". Geobios. 62: 23–29. doi:10.1016/j.geobios.2020.07.003.
- Li, Rui-Yun; Wang, Xue-lian; Chen, Jing-Wei; Deng, Sheng-Hui; Wang, Zi-Xi; Dong, Jun-Ling; Sun, Bai-Nian (June 2016). "A new thalloid liverwort: Pallaviciniites sandaolingensis sp. nov. from the Middle Jurassic of Turpan–Hami Basin, NW China". PalZ. 90 (2): 389–397. doi:10.1007/s12542-016-0299-3. ISSN 0031-0220. S2CID 131295547.
- Li, Ruiyun; Li, Xiaoqiang; Wang, Hongshan; Sun, Bainian (2019). "Ricciopsis sandaolingensis sp. nov., a new fossil bryophyte from the Middle Jurassic Xishanyao Formation in the Turpan-Hami Basin, Xinjiang, Northwest China". Palaeontologia Electronica. 22 (2). doi:10.26879/917. ISSN 1094-8074.
- Na, Yuling; Sun, Chunlin; Wang, Hongshan; Dilcher, David L.; Li, Yunfeng; Li, Tao (December 2017). "A brief introduction to the Middle Jurassic Daohugou Flora from Inner Mongolia, China". Review of Palaeobotany and Palynology. 247: 53–67. doi:10.1016/j.revpalbo.2017.08.003.
- Pole, Mike (June 2009). "Vegetation and climate of the New Zealand Jurassic". GFF. 131 (1–2): 105–111. doi:10.1080/11035890902808948. ISSN 1103-5897. S2CID 140686834.
- Cleal, C. J.; Rees, P. M. (July 2003). "The Middle Jurassic flora from Stonesfield, Oxfordshire, UK". Palaeontology. 46 (4): 739–801. doi:10.1111/1475-4983.00319. ISSN 0031-0239.
- Motani 2000.
- Vinn & Mutvei 2009, pp. 286-296.
- Vinn, ten Hove & Mutvei 2008, pp. 295–301.
- Taylor & Wilson 2003, pp. 1-103.
- Haines 2000.
- Feduccia 1996.
- Witton, Martill & Loveridge 2010, pp. 79-81.
- Witton 2016.
- Carroll 1988.
- Toljagić, Olja; Butler, Richard J. (2013-06-23). "Triassic–Jurassic mass extinction as trigger for the Mesozoic radiation of crocodylomorphs". Biology Letters. 9 (3): 20130095. doi:10.1098/rsbl.2013.0095. ISSN 1744-9561. PMC 3645043. PMID 23536443.
- Melstrom, Keegan M.; Irmis, Randall B. (July 2019). "Repeated Evolution of Herbivorous Crocodyliforms during the Age of Dinosaurs". Current Biology. 29 (14): 2389–2395.e3. doi:10.1016/j.cub.2019.05.076. PMID 31257139. S2CID 195699188.
- Irmis, Randall B.; Nesbitt, Sterling J.; Sues, Hans-Dieter (2013). "Early Crocodylomorpha". Geological Society, London, Special Publications. 379 (1): 275–302. Bibcode:2013GSLSP.379..275I. doi:10.1144/SP379.24. ISSN 0305-8719. S2CID 219190410.
- Leardi, Juan Martin; Pol, Diego; Clark, James Matthew (2017-01-19). "Detailed anatomy of the braincase of Macelognathus vagans Marsh, 1884 (Archosauria, Crocodylomorpha) using high resolution tomography and new insights on basal crocodylomorph phylogeny". PeerJ. 5: e2801. doi:10.7717/peerj.2801. ISSN 2167-8359. PMC 5251941. PMID 28133565.
- Wilberg, Eric W.; Turner, Alan H.; Brochu, Christopher A. (2019-01-24). "Evolutionary structure and timing of major habitat shifts in Crocodylomorpha". Scientific Reports. 9 (1): 514. Bibcode:2019NatSR...9..514W. doi:10.1038/s41598-018-36795-1. ISSN 2045-2322. PMC 6346023. PMID 30679529.
- Dal Sasso, C.; Pasini, G.; Fleury, G.; Maganuco, S. (2017). "Razanandrongobe sakalavae, a gigantic mesoeucrocodylian from the Middle Jurassic of Madagascar, is the oldest known notosuchian". PeerJ. 5: e3481. doi:10.7717/peerj.3481. PMC 5499610. PMID 28690926.
- Joyce, Walter G. (April 2017). "A Review of the Fossil Record of Basal Mesozoic Turtles". Bulletin of the Peabody Museum of Natural History. 58 (1): 65–113. doi:10.3374/014.058.0105. ISSN 0079-032X. S2CID 54982901.
- Sterli, Juliana; de la Fuente, Marcelo S.; Rougier, Guillermo W. (2018-07-04). "New remains of Condorchelys antiqua (Testudinata) from the Early-Middle Jurassic of Patagonia: anatomy, phylogeny, and paedomorphosis in the early evolution of turtles". Journal of Vertebrate Paleontology. 38 (4): (1)–(17). doi:10.1080/02724634.2018.1480112. ISSN 0272-4634. S2CID 109556104.
- Wings et al. 2012, pp. 925-935.
- Gannon 2012.
- Rabi, Márton; Joyce, Walter G.; Wings, Oliver (September 2010). "A review of the Mesozoic turtles of the Junggar Basin (Xinjiang, Northwest China) and the paleobiogeography of Jurassic to Early Cretaceous Asian testudinates". Palaeobiodiversity and Palaeoenvironments. 90 (3): 259–273. doi:10.1007/s12549-010-0031-3. ISSN 1867-1594. S2CID 128805609.
- Sullivan, Patrick M.; Joyce, Walter G. (August 2017). "The shell and pelvic anatomy of the Late Jurassic turtle Platychelys oberndorferi based on material from Solothurn, Switzerland". Swiss Journal of Palaeontology. 136 (2): 323–343. doi:10.1007/s13358-017-0136-7. ISSN 1664-2376. S2CID 90587841.
- Anquetin, Jérémy; Püntener, Christian; Joyce, Walter G. (October 2017). "A Review of the Fossil Record of Turtles of the Clade Thalassochelydia". Bulletin of the Peabody Museum of Natural History. 58 (2): 317–369. doi:10.3374/014.058.0205. ISSN 0079-032X. S2CID 31091127.
- Cleary, Terri J.; Benson, Roger B. J.; Evans, Susan E.; Barrett, Paul M. (21 March 2018). "Lepidosaurian diversity in the Mesozoic–Palaeogene: the potential roles of sampling biases and environmental drivers". Royal Society Open Science. 5 (3): 171830. doi:10.1098/rsos.171830. PMC 5882712. PMID 29657788.
- Herrera‐Flores, Jorge A.; Stubbs, Thomas L.; Benton, Michael J. (2017). "Macroevolutionary patterns in Rhynchocephalia: is the tuatara (Sphenodon punctatus) a living fossil?". Palaeontology. 60 (3): 319–328. doi:10.1111/pala.12284. ISSN 1475-4983.
- Evans, Susan E.; Jones, Marc E.H. (2010), "The Origin, Early History and Diversification of Lepidosauromorph Reptiles", New Aspects of Mesozoic Biodiversity, Berlin, Heidelberg: Springer Berlin Heidelberg, 132, pp. 27–44, doi:10.1007/978-3-642-10311-7_2, ISBN 978-3-642-10310-0, retrieved 2021-01-07
- Burbrink, Frank T; Grazziotin, Felipe G; Pyron, R Alexander; Cundall, David; Donnellan, Steve; Irish, Frances; Keogh, J Scott; Kraus, Fred; Murphy, Robert W; Noonan, Brice; Raxworthy, Christopher J (2020-05-01). Thomson, Robert (ed.). "Interrogating Genomic-Scale Data for Squamata (Lizards, Snakes, and Amphisbaenians) Shows no Support for Key Traditional Morphological Relationships". Systematic Biology. 69 (3): 502–520. doi:10.1093/sysbio/syz062. ISSN 1063-5157. PMID 31550008.
- Dong, Liping; Wang, Yuan; Mou, Lijie; Zhang, Guoze; Evans, Susan E. (2019-09-13). "A new Jurassic lizard from China". Geodiversitas. 41 (16): 623. doi:10.5252/geodiversitas2019v41a16. ISSN 1280-9659. S2CID 204256127.
- Caldwell, M. W.; Nydam, R. L.; Palci, A.; Apesteguía, S. N. (2015). "The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution". Nature Communications. 6: 5996. Bibcode:2015NatCo...6.5996C. doi:10.1038/ncomms6996. PMID 25625704.
- Evans, Susan E.; Chure, Daniel C. (1998-04-10). "Paramacellodid lizard skulls from the Jurassic Morrison Formation at Dinosaur National Monument, Utah". Journal of Vertebrate Paleontology. 18 (1): 99–114. doi:10.1080/02724634.1998.10011037. ISSN 0272-4634.
- Simões, Tiago R.; Caldwell, Michael W.; Nydam, Randall L.; Jiménez-Huidobro, Paulina (September 2016). "Osteology, phylogeny, and functional morphology of two Jurassic lizard species and the early evolution of scansoriality in geckoes". Zoological Journal of the Linnean Society. doi:10.1111/zoj.12487.
- Daza, J. D.; Bauer, A. M.; Stanley, E. L.; Bolet, A.; Dickson, B.; Losos, J. B. (2018-11-01). "An Enigmatic Miniaturized and Attenuate Whole Lizard from the Mid-Cretaceous Amber of Myanmar". Breviora. 563 (1): 1. doi:10.3099/MCZ49.1. ISSN 0006-9698. S2CID 91589111.
- Evans, S. E. (1991). "A new lizard-like reptile (Diapsida: Lepidosauromorpha) from the Middle Jurassic of England". Zoological Journal of the Linnean Society. 103 (4): 391–412. doi:10.1111/j.1096-3642.1991.tb00910.x.
- Matsumoto, R.; Evans, S. E. (2010). "Choristoderes and the freshwater assemblages of Laurasia". Journal of Iberian Geology. 36 (2): 253–274. doi:10.5209/rev_JIGE.2010.v36.n2.11. ISSN 1698-6180.
- Matsumoto, Ryoko; Dong, Liping; Wang, Yuan; Evans, Susan E. (2019-06-18). "The first record of a nearly complete choristodere (Reptilia: Diapsida) from the Upper Jurassic of Hebei Province, People's Republic of China". Journal of Systematic Palaeontology. 17 (12): 1031–1048. doi:10.1080/14772019.2018.1494220. ISSN 1477-2019. S2CID 92421503.
- Moon, Benjamin C.; Stubbs, Thomas L. (2020-02-13). "Early high rates and disparity in the evolution of ichthyosaurs". Communications Biology. 3 (1): 68. doi:10.1038/s42003-020-0779-6. ISSN 2399-3642. PMC 7018711. PMID 32054967.
- Thorne, P. M.; Ruta, M.; Benton, M. J. (2011-05-17). "Resetting the evolution of marine reptiles at the Triassic-Jurassic boundary". Proceedings of the National Academy of Sciences. 108 (20): 8339–8344. Bibcode:2011PNAS..108.8339T. doi:10.1073/pnas.1018959108. ISSN 0027-8424. PMC 3100925. PMID 21536898.
- Druckenmiller, Patrick S.; Maxwell, Erin E. (January 2014). "A Middle Jurassic (Bajocian) ophthalmosaurid (Reptilia, Ichthyosauria) from the Tuxedni Formation, Alaska and the early diversification of the clade". Geological Magazine. 151 (1): 41–48. Bibcode:2014GeoM..151...41D. doi:10.1017/S0016756813000125. ISSN 0016-7568.
- Wintrich, Tanja; Hayashi, Shoji; Houssaye, Alexandra; Nakajima, Yasuhisa; Sander, P. Martin (2017-12-01). "A Triassic plesiosaurian skeleton and bone histology inform on evolution of a unique body plan". Science Advances. 3 (12): e1701144. Bibcode:2017SciA....3E1144W. doi:10.1126/sciadv.1701144. ISSN 2375-2548. PMC 5729018. PMID 29242826.
- Benson, Roger B. J.; Evans, Mark; Druckenmiller, Patrick S. (2012-03-16). "High Diversity, Low Disparity and Small Body Size in Plesiosaurs (Reptilia, Sauropterygia) from the Triassic–Jurassic Boundary". PLOS ONE. 7 (3): e31838. Bibcode:2012PLoSO...731838B. doi:10.1371/journal.pone.0031838. ISSN 1932-6203. PMC 3306369. PMID 22438869.
- O'Keefe, F. Robin (2002). "The evolution of plesiosaur and pliosaur morphotypes in the Plesiosauria (Reptilia: Sauropterygia)". Paleobiology. 28 (1): 101–112. doi:10.1666/0094-8373(2002)028<0101:TEOPAP>2.0.CO;2. ISSN 0094-8373.
- Benson, Roger B. J.; Evans, Mark; Smith, Adam S.; Sassoon, Judyth; Moore-Faye, Scott; Ketchum, Hilary F.; Forrest, Richard (2013-05-31). "A Giant Pliosaurid Skull from the Late Jurassic of England". PLOS ONE. 8 (5): e65989. Bibcode:2013PLoSO...865989B. doi:10.1371/journal.pone.0065989. ISSN 1932-6203. PMC 3669260. PMID 23741520.
- Ketchum, Hilary F.; Benson, Roger B. J. (12 April 2010). "Global interrelationships of Plesiosauria (Reptilia, Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses". Biological Reviews. 85 (2): 361–392. doi:10.1111/j.1469-185X.2009.00107.x. PMID 20002391. S2CID 12193439.
- Gao, Ting; Li, Da-Qing; Li, Long-Feng; Yang, Jing-Tao (2019-08-13). "The first record of freshwater plesiosaurian from the Middle Jurassic of Gansu, NW China, with its implications to the local palaeobiogeography". Journal of Palaeogeography. 8 (1): 27. Bibcode:2019JPalg...8...27G. doi:10.1186/s42501-019-0043-5. ISSN 2524-4507. S2CID 199547716.
- Kear, Benjamin P. (2 August 2012). "A revision of Australia's Jurassic plesiosaurs". Palaeontology. 55 (5): 1125–1138. doi:10.1111/j.1475-4983.2012.01183.x.
- O’Sullivan, Michael; Martill, David M. (2017-11-17). "The taxonomy and systematics of Parapsicephalus purdoni (Reptilia: Pterosauria) from the Lower Jurassic Whitby Mudstone Formation, Whitby, U.K". Historical Biology. 29 (8): 1009–1018. doi:10.1080/08912963.2017.1281919. ISSN 0891-2963. S2CID 132532024.
- Bestwick, Jordan; Unwin, David M.; Butler, Richard J.; Henderson, Donald M.; Purnell, Mark A. (November 2018). "Pterosaur dietary hypotheses: a review of ideas and approaches: Pterosaur dietary hypotheses". Biological Reviews. 93 (4): 2021–2048. doi:10.1111/brv.12431. PMC 6849529. PMID 29877021.
- Brusatte, Stephen L; Benton, Michael J; Ruta, Marcello; Lloyd, Graeme T (2008-12-23). "The first 50 Myr of dinosaur evolution: macroevolutionary pattern and morphological disparity". Biology Letters. 4 (6): 733–736. doi:10.1098/rsbl.2008.0441. PMC 2614175. PMID 18812311.
- Temp Müller, Rodrigo; Augusto Pretto, Flávio; Kerber, Leonardo; Silva-Neves, Eduardo; Dias-da-Silva, Sérgio (28 March 2018). "Comment on 'A dinosaur missing-link? Chilesaurus and the early evolution of ornithischian dinosaurs'". Biology Letters. 14 (3): 20170581. doi:10.1098/rsbl.2017.0581. ISSN 1744-9561. PMC 5897605. PMID 29593074.
- Zahner, Marion; Brinkmann, Winand (August 2019). "A Triassic averostran-line theropod from Switzerland and the early evolution of dinosaurs". Nature Ecology & Evolution. 3 (8): 1146–1152. doi:10.1038/s41559-019-0941-z. ISSN 2397-334X. PMC 6669044. PMID 31285577.
- Sasso, Cristiano Dal; Maganuco, Simone; Cau, Andrea (2018-12-19). "The oldest ceratosaurian (Dinosauria: Theropoda), from the Lower Jurassic of Italy, sheds light on the evolution of the three-fingered hand of birds". PeerJ. 6: e5976. doi:10.7717/peerj.5976. ISSN 2167-8359. PMC 6304160. PMID 30588396.
- Wang, Shuo; Stiegler, Josef; Amiot, Romain; Wang, Xu; Du, Guo-hao; Clark, James M.; Xu, Xing (January 2017). "Extreme Ontogenetic Changes in a Ceratosaurian Theropod". Current Biology. 27 (1): 144–148. doi:10.1016/j.cub.2016.10.043. PMID 28017609. S2CID 441498.
- Zanno, Lindsay E.; Makovicky, Peter J. (2011-01-04). "Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution". Proceedings of the National Academy of Sciences. 108 (1): 232–237. Bibcode:2011PNAS..108..232Z. doi:10.1073/pnas.1011924108. ISSN 0027-8424. PMC 3017133. PMID 21173263.
- Rauhut, Oliver W. M.; Foth, Christian (2020), Foth, Christian; Rauhut, Oliver W. M. (eds.), "The Origin of Birds: Current Consensus, Controversy, and the Occurrence of Feathers", The Evolution of Feathers: From Their Origin to the Present, Fascinating Life Sciences, Cham: Springer International Publishing, pp. 27–45, doi:10.1007/978-3-030-27223-4_3, ISBN 978-3-030-27223-4, retrieved 2021-01-05
- Benson, R.B.J (2010). "A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods". Zoological Journal of the Linnean Society. 158 (4): 882–935. doi:10.1111/j.1096-3642.2009.00569.x.
- Rauhut, Oliver W. M.; Pol, Diego (2019-12-11). "Probable basal allosauroid from the early Middle Jurassic Cañadón Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs". Scientific Reports. 9 (1): 18826. doi:10.1038/s41598-019-53672-7. ISSN 2045-2322. PMC 6906444. PMID 31827108.
- Rauhut, Oliver W. M.; Milner, Angela C.; Moore-Fay, Scott (2010). "Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi(Woodward, 1910) from the Middle Jurassic of England". Zoological Journal of the Linnean Society. 158: 155–195. doi:10.1111/j.1096-3642.2009.00591.x.
- Qin, Z., Clark, J., Choiniere, J., & Xu, X. (2019). A new alvarezsaurian theropod from the Upper Jurassic Shishugou Formation of western China. Scientific Reports, 9: 11727. doi:10.1038/s41598-019-48148-7
- Wang, Min; O’Connor, Jingmai K.; Xu, Xing; Zhou, Zhonghe (May 2019). "A new Jurassic scansoriopterygid and the loss of membranous wings in theropod dinosaurs". Nature. 569 (7755): 256–259. Bibcode:2019Natur.569..256W. doi:10.1038/s41586-019-1137-z. ISSN 1476-4687. PMID 31068719. S2CID 148571099.
- Hartman, Scott; Mortimer, Mickey; Wahl, William R.; Lomax, Dean R.; Lippincott, Jessica; Lovelace, David M. (2019-07-10). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247. doi:10.7717/peerj.7247. ISSN 2167-8359. PMC 6626525. PMID 31333906.
- Norman, David B (2021-01-01). "Scelidosaurus harrisonii (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: biology and phylogenetic relationships". Zoological Journal of the Linnean Society. 191 (1): 1–86. doi:10.1093/zoolinnean/zlaa061. ISSN 0024-4082.
- Godefroit, Pascal; Sinitsa, Sofia M.; Cincotta, Aude; McNamara, Maria E.; Reshetova, Svetlana A.; Dhouailly, Danielle (2020), Foth, Christian; Rauhut, Oliver W. M. (eds.), "Integumentary Structures in Kulindadromeus zabaikalicus, a Basal Neornithischian Dinosaur from the Jurassic of Siberia", The Evolution of Feathers: From Their Origin to the Present, Fascinating Life Sciences, Cham: Springer International Publishing, pp. 47–65, doi:10.1007/978-3-030-27223-4_4, ISBN 978-3-030-27223-4, retrieved 2021-01-05
- McDonald, Andrew T. (2012-05-22). Farke, Andrew A. (ed.). "Phylogeny of Basal Iguanodonts (Dinosauria: Ornithischia): An Update". PLOS ONE. 7 (5): e36745. Bibcode:2012PLoSO...736745M. doi:10.1371/journal.pone.0036745. ISSN 1932-6203. PMC 3358318. PMID 22629328.
- Han, Fenglu; Forster, Catherine A.; Clark, James M.; Xu, Xing (2015-12-09). "A New Taxon of Basal Ceratopsian from China and the Early Evolution of Ceratopsia". PLOS ONE. 10 (12): e0143369. Bibcode:2015PLoSO..1043369H. doi:10.1371/journal.pone.0143369. ISSN 1932-6203. PMC 4674058. PMID 26649770.
- McPhee, Blair W.; Benson, Roger B.J.; Botha-Brink, Jennifer; Bordy, Emese M.; Choiniere, Jonah N. (8 October 2018). "A Giant Dinosaur from the Earliest Jurassic of South Africa and the Transition to Quadrupedality in Early Sauropodomorphs". Current Biology. 28 (19): 3143–3151.e7. doi:10.1016/j.cub.2018.07.063. PMID 30270189. S2CID 52890502.
- Viglietti, Pia A.; Barrett, Paul M.; Broderick, Tim J.; Munyikwa, Darlington; MacNiven, Rowan; Broderick, Lucy; Chapelle, Kimberley; Glynn, Dave; Edwards, Steve; Zondo, Michel; Broderick, Patricia (January 2018). "Stratigraphy of the Vulcanodon type locality and its implications for regional correlations within the Karoo Supergroup". Journal of African Earth Sciences. 137: 149–156. Bibcode:2018JAfES.137..149V. doi:10.1016/j.jafrearsci.2017.10.015.
- Royo-Torres, Rafael; Cobos, Alberto; Mocho, Pedro; Alcalá, Luis (2021-01-01). "Origin and evolution of turiasaur dinosaurs set by means of a new 'rosetta' specimen from Spain". Zoological Journal of the Linnean Society. 191 (1): 201–227. doi:10.1093/zoolinnean/zlaa091. ISSN 0024-4082.
- Ren, Xin-Xin; Huang, Jian-Dong; You, Hai-Lu (2020-05-27). "The second mamenchisaurid dinosaur from the Middle Jurassic of Eastern China". Historical Biology. 32 (5): 602–610. doi:10.1080/08912963.2018.1515935. ISSN 0891-2963. S2CID 91927243.
- Xu, Xing; Upchurch, Paul; Mannion, Philip D.; Barrett, Paul M.; Regalado-Fernandez, Omar R.; Mo, Jinyou; Ma, Jinfu; Liu, Hongan (2018-07-24). "A new Middle Jurassic diplodocoid suggests an earlier dispersal and diversification of sauropod dinosaurs". Nature Communications. 9 (1): 2700. Bibcode:2018NatCo...9.2700X. doi:10.1038/s41467-018-05128-1. ISSN 2041-1723. PMC 6057878. PMID 30042444.
- Philip D. Mannion, Ronan Allain & Olivier Moine, 2017, "The earliest known titanosauriform sauropod dinosaur and the evolution of Brachiosauridae", PeerJ 5: e3217 https://doi.org/10.7717/peerj.3217
- Tschopp, Emanuel; Mateus, Octávio; Benson, Roger B. J. (2015-04-07). "A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)". PeerJ. 3: e857. doi:10.7717/peerj.857. ISSN 2167-8359. PMC 4393826. PMID 25870766.
- Sander, P. Martin; Christian, Andreas; Clauss, Marcus; Fechner, Regina; Gee, Carole T.; Griebeler, Eva-Maria; Gunga, Hanns-Christian; Hummel, Jürgen; Mallison, Heinrich; Perry, Steven F.; Preuschoft, Holger (February 2011). "Biology of the sauropod dinosaurs: the evolution of gigantism". Biological Reviews. 86 (1): 117–155. doi:10.1111/j.1469-185X.2010.00137.x. PMC 3045712. PMID 21251189.
- Averianov, Alexander O.; Martin, Thomas; Skutschas, Pavel P.; Rezvyi, Anton S.; Bakirov, Aizek A. (March 2008). "Amphibians from the Middle Jurassic Balabansai Svita in the Fergana Depression, Kyrgyzstan (Central Asia)". Palaeontology. 51 (2): 471–485. doi:10.1111/j.1475-4983.2007.00748.x.
- Warren, A. A.; Hutchinson, M. N.; Hill, Dorothy (1983-09-13). "The last Labyrinthodont? A new brachyopoid (Amphibia, Temnospondyli) from the early Jurassic Evergreen formation of Queensland, Australia". Philosophical Transactions of the Royal Society of London. B, Biological Sciences. 303 (1113): 1–62. Bibcode:1983RSPTB.303....1W. doi:10.1098/rstb.1983.0080.
- Reilly, Stephen M.; Jorgensen, Michael E. (February 2011). "The evolution of jumping in frogs: Morphological evidence for the basal anuran locomotor condition and the radiation of locomotor systems in crown group anurans". Journal of Morphology. 272 (2): 149–168. doi:10.1002/jmor.10902. PMID 21210487. S2CID 14217777.
- Báez, Ana Maria; Nicoli, Laura (March 2008). "A new species of Notobatrachus (Amphibia, Salientia) from the Middle Jurassic of northwestern Patagonia". Journal of Paleontology. 82 (2): 372–376. doi:10.1666/06-117.1. ISSN 0022-3360. S2CID 130032431.
- Schoch, Rainer R.; Werneburg, Ralf; Voigt, Sebastian (2020-05-26). "A Triassic stem-salamander from Kyrgyzstan and the origin of salamanders". Proceedings of the National Academy of Sciences. 117 (21): 11584–11588. doi:10.1073/pnas.2001424117. ISSN 0027-8424. PMC 7261083. PMID 32393623.
- Skutschas, Pavel; Stein, Koen (April 2015). "Long bone histology of the stem salamander Kokartus honorarius (Amphibia: Caudata) from the Middle Jurassic of Kyrgyzstan". Journal of Anatomy. 226 (4): 334–347. doi:10.1111/joa.12281. PMC 4386933. PMID 25682890.
- Jia, Jia; Gao, Ke-Qin (2019-03-04). "A new stem hynobiid salamander (Urodela, Cryptobranchoidea) from the Upper Jurassic (Oxfordian) of Liaoning Province, China". Journal of Vertebrate Paleontology. 39 (2): e1588285. doi:10.1080/02724634.2019.1588285. ISSN 0272-4634. S2CID 164310171.
- Gao, K.-Q.; Shubin, N. H. (2012-04-10). "Late Jurassic salamandroid from western Liaoning, China". Proceedings of the National Academy of Sciences. 109 (15): 5767–5772. doi:10.1073/pnas.1009828109. ISSN 0027-8424. PMC 3326464. PMID 22411790.
- Evans, S. E.; Lally, C.; Chure, D. C.; Elder, A.; Maisano, J. A. (2005). "A Late Jurassic salamander (Amphibia: Caudata) from the Morrison Formation of North America". Zoological Journal of the Linnean Society. 143 (4): 599–616. doi:10.1111/j.1096-3642.2005.00159.x.
- Santos, Rodolfo Otávio; Laurin, Michel; Zaher, Hussam (2020-11-03). "A review of the fossil record of caecilians (Lissamphibia: Gymnophionomorpha) with comments on its use to calibrate molecular timetrees". Biological Journal of the Linnean Society. 131 (4): 737–755. doi:10.1093/biolinnean/blaa148. ISSN 0024-4066.
- Haddoumi, Hamid; Allain, Ronan; Meslouh, Said; Metais, Grégoire; Monbaron, Michel; Pons, Denise; Rage, Jean-Claude; Vullo, Romain; Zouhri, Samir (January 2016). "Guelb el Ahmar (Bathonian, Anoual Syncline, eastern Morocco): First continental flora and fauna including mammals from the Middle Jurassic of Africa" (PDF). Gondwana Research. 29 (1): 290–319. Bibcode:2016GondR..29..290H. doi:10.1016/j.gr.2014.12.004. ISSN 1342-937X.
- Lee, Michael S.Y.; Beck, Robin M.D. (31 August 2015). "Mammalian Evolution: A Jurassic Spark". Current Biology. 25 (17): R759–R761. doi:10.1016/j.cub.2015.07.008. PMID 26325137. S2CID 11088107.
- Ji, Q.; Luo, Z.-X.; Yuan, C.-X.; Tabrum, A. R. (2006). "A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals". Science. 311 (5, 764): 1, 123–1, 127. Bibcode:2006Sci...311.1123J. doi:10.1126/science.1123026. PMID 16497926. S2CID 46067702.
- Meng, Qing-Jin; Grossnickle, David M.; Liu, Di; Zhang, Yu-Guang; Neander, April I.; Ji, Qiang; Luo, Zhe-Xi (August 2017). "New gliding mammaliaforms from the Jurassic". Nature. 548 (7667): 291–296. Bibcode:2017Natur.548..291M. doi:10.1038/nature23476. ISSN 1476-4687. PMID 28792929. S2CID 205259206.
- Meng, J.; Hu, Y.-M.; Wang, Y.-Q.; Wang, X.-L.; Li, C.-K. (2007). "Corrigendum: A Mesozoic gliding mammal from northeastern China". Nature 446 (7131): 102. Bibcode:2007Natur.446Q.102M. doi:10.1038/nature05639.
- Luo, Z.-X.; Wible, J.R. (2005). "A Late Jurassic Digging Mammal and Early Mammalian Diversification". Science. 308 (5718): 103–107. Bibcode:2005Sci...308..103L. doi:10.1126/science.1108875. ISSN 0036-8075. PMID 15802602. S2CID 7031381.
- Luo, Zhe-Xi; Cifelli, Richard L.; Kielan-Jaworowska, Zofia (January 2001). "Dual origin of tribosphenic mammals". Nature. 409 (6816): 53–57. Bibcode:2001Natur.409...53L. doi:10.1038/35051023. ISSN 0028-0836. PMID 11343108. S2CID 4342585.
- Zhe-Xi Luo; Chong-Xi Yuan; Qing-Jin Meng; Qiang Ji (25 August 2011). "A Jurassic eutherian mammal and divergence of marsupials and placentals" (PDF). Nature. 476 (7361): 442–445. Bibcode:2011Natur.476..442L. doi:10.1038/nature10291. PMID 21866158. S2CID 205225806. Archived from the original (PDF) on 10 November 2013. Electronic supplementary material
- King, Benedict; Beck, Robin M. D. (2020-06-10). "Tip dating supports novel resolutions of controversial relationships among early mammals". Proceedings of the Royal Society B: Biological Sciences. 287 (1928): 20200943. doi:10.1098/rspb.2020.0943. PMC 7341916. PMID 32517606.
- Ruta, Marcello; Botha-Brink, Jennifer; Mitchell, Stephen A.; Benton, Michael J. (2013-10-22). "The radiation of cynodonts and the ground plan of mammalian morphological diversity". Proceedings of the Royal Society B: Biological Sciences. 280 (1769): 20131865. doi:10.1098/rspb.2013.1865. ISSN 0962-8452. PMC 3768321. PMID 23986112.
- Du, Yixing; Chiari, Marco; Karádi, Viktor; Nicora, Alda; Onoue, Tetsuji; Pálfy, József; Roghi, Guido; Tomimatsu, Yuki; Rigo, Manuel (April 2020). "The asynchronous disappearance of conodonts: New constraints from Triassic-Jurassic boundary sections in the Tethys and Panthalassa". Earth-Science Reviews. 203: 103176. Bibcode:2020ESRv..20303176D. doi:10.1016/j.earscirev.2020.103176.
- Ginot, Samuel; Goudemand, Nicolas (December 2020). "Global climate changes account for the main trends of conodont diversity but not for their final demise". Global and Planetary Change. 195: 103325. Bibcode:2020GPC...19503325G. doi:10.1016/j.gloplacha.2020.103325.
- Kemp, Anne; Cavin, Lionel; Guinot, Guillaume (April 2017). "Evolutionary history of lungfishes with a new phylogeny of post-Devonian genera". Palaeogeography, Palaeoclimatology, Palaeoecology. 471: 209–219. Bibcode:2017PPP...471..209K. doi:10.1016/j.palaeo.2016.12.051.
- Cavin, Lionel; Cupello, Camila; Yabumoto, Yoshitaka; Léo, Fragoso; Deersi, Uthumporn; Brito, Paul M. (2019). "Phylogeny and evolutionary history of mawsoniid coelacanths" (PDF). Bulletin of the Kitakyushu Museum of Natural History and Human History, Series A. 17: 3–13.
- Clement, Gaël (2005-09-30). "A new coelacanth (Actinistia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria". Journal of Vertebrate Paleontology. 25 (3): 481–491. doi:10.1671/0272-4634(2005)025[0481:ANCASF]2.0.CO;2. ISSN 0272-4634.
- Poyato-Ariza, Francisco José; Martín-Abad, Hugo (2020-07-19). "History of two lineages: Comparative analysis of the fossil record in Amiiformes and Pycnodontiformes (Osteischtyes, Actinopterygii)". Spanish Journal of Palaeontology. 28 (1): 79. doi:10.7203/sjp.28.1.17833. ISSN 2255-0550.
- Arratia G. Mesozoic halecostomes and the early radiation of teleosts. In: Arratia G, Tintori A, editors. Mesozoic Fishes 3 – Systematics, Paleoenvironments and Biodiversity. München: Verlag Dr. Friedrich Pfeil; 2004. p. 279–315.
- Tse, Tze-Kei; Pittman, Michael; Chang, Mee-mann (2015-03-26). "A specimen of Paralycoptera Chang & Chou 1977 (Teleostei: Osteoglossoidei) from Hong Kong (China) with a potential Late Jurassic age that extends the temporal and geographical range of the genus". PeerJ. 3: e865. doi:10.7717/peerj.865. ISSN 2167-8359. PMC 4380157. PMID 25834774.
- Liston, J., Newbrey, M., Challands, T., and Adams, C., 2013, "Growth, age and size of the Jurassic pachycormid Leedsichthys problematicus (Osteichthyes: Actinopterygii) in: Arratia, G., Schultze, H. and Wilson, M. (eds.) Mesozoic Fishes 5 – Global Diversity and Evolution. Verlag Dr. Friedrich Pfeil, München, Germany, pp. 145–175
- Rees, Jan; Underwood, Charlie J. (2008-01-17). "Hybodont sharks of the English Bathonian and Callovian (Middle Jurassic)". Palaeontology. 51 (1): 117–147. doi:10.1111/j.1475-4983.2007.00737.x.
- Underwood, Charlie J. (March 2006). "Diversification of the Neoselachii (Chondrichthyes) during the Jurassic and Cretaceous". Paleobiology. 32 (2): 215–235. doi:10.1666/04069.1. ISSN 0094-8373. S2CID 86232401.
- Kriwet, Jürgen; Klug, Stefanie (December 2011). "A new Jurassic cow shark (Chondrichthyes, Hexanchiformes) with comments on Jurassic hexanchiform systematics". Swiss Journal of Geosciences. 104 (S1): 107–114. doi:10.1007/s00015-011-0075-z. ISSN 1661-8726. S2CID 84405176.
- Stumpf, Sebastian; Kriwet, Jürgen (2019-12-01). "A new Pliensbachian elasmobranch (Vertebrata, Chondrichthyes) assemblage from Europe, and its contribution to the understanding of late Early Jurassic elasmobranch diversity and distributional patterns". PalZ. 93 (4): 637–658. doi:10.1007/s12542-019-00451-4. ISSN 1867-6812. S2CID 181782998.
- Underwood, Charlie J.; Claeson, Kerin M. (June 2019). "The Late Jurassic ray Kimmerobatis etchesi gen. et sp. nov. and the Jurassic radiation of the Batoidea". Proceedings of the Geologists' Association. 130 (3–4): 345–354. doi:10.1016/j.pgeola.2017.06.009.
- Slater, Tiffany S.; Ashbrook, Kate; Kriwet, Jürgen (August 2020). Cavin, Lionel (ed.). "Evolutionary relationships among bullhead sharks (Chondrichthyes, Heterodontiformes)". Papers in Palaeontology. 6 (3): 425–437. doi:10.1002/spp2.1299. ISSN 2056-2802.
- Srdic, Alex; Duffin, Christopher J.; Martill, David M. (August 2016). "First occurrence of the orectolobiform shark Akaimia in the Oxford Clay Formation (Jurassic, Callovian) of England". Proceedings of the Geologists' Association. 127 (4): 506–513. doi:10.1016/j.pgeola.2016.07.002.
- Jambura, Patrick L.; Kindlimann, René; López-Romero, Faviel; Marramà, Giuseppe; Pfaff, Cathrin; Stumpf, Sebastian; Türtscher, Julia; Underwood, Charlie J.; Ward, David J.; Kriwet, Jürgen (December 2019). "Micro-computed tomography imaging reveals the development of a unique tooth mineralization pattern in mackerel sharks (Chondrichthyes; Lamniformes) in deep time". Scientific Reports. 9 (1): 9652. Bibcode:2019NatSR...9.9652J. doi:10.1038/s41598-019-46081-3. ISSN 2045-2322. PMC 6609643. PMID 31273249.
- López-Romero, Faviel A.; Stumpf, Sebastian; Pfaff, Cathrin; Marramà, Giuseppe; Johanson, Zerina; Kriwet, Jürgen (2020-07-28). "Evolutionary trends of the conserved neurocranium shape in angel sharks (Squatiniformes, Elasmobranchii)". Scientific Reports. 10 (1): 12582. Bibcode:2020NatSR..1012582L. doi:10.1038/s41598-020-69525-7. ISSN 2045-2322. PMC 7387474. PMID 32724124.
- Stumpf, Sebastian; Scheer, Udo; Kriwet, Jürgen (2019-03-04). "A new genus and species of extinct ground shark, †Diprosopovenator hilperti, gen. et sp. nov. (Carcharhiniformes, †Pseudoscyliorhinidae, fam. nov.), from the Upper Cretaceous of Germany". Journal of Vertebrate Paleontology. 39 (2): e1593185. doi:10.1080/02724634.2019.1593185. ISSN 0272-4634. S2CID 155785248.
- Popov, Evgeny V.; Delsate, Dominique; Felten, Roland (2019-07-02). "A New Callorhinchid Genus (Holocephali, Chimaeroidei) from the Early Bajocian of Ottange-Rumelange, on the Luxembourg-French Border". Paleontological Research. 23 (3): 220. doi:10.2517/2018PR021. ISSN 1342-8144.
- Duffin, Christopher J.; Milàn, Jesper (2017-11-14). "A new myriacanthid holocephalian from the Early Jurassic of Denmark". Bulletin of the Geological Society of Denmark. 65: 161–170. doi:10.37570/bgsd-2017-65-10. ISSN 2245-7070.
- Labandeira, Conrad C. (2018-05-23), "The Fossil History of Insect Diversity", Insect Biodiversity, Chichester, UK: John Wiley & Sons, Ltd, pp. 723–788, doi:10.1002/9781118945582.ch24, ISBN 978-1-118-94558-2, retrieved 2020-12-06
- McKenna, Duane D.; Shin, Seunggwan; Ahrens, Dirk; Balke, Michael; Beza-Beza, Cristian; Clarke, Dave J.; Donath, Alexander; Escalona, Hermes E.; Friedrich, Frank; Letsch, Harald; Liu, Shanlin (2019-12-03). "The evolution and genomic basis of beetle diversity". Proceedings of the National Academy of Sciences. 116 (49): 24729–24737. doi:10.1073/pnas.1909655116. ISSN 0027-8424. PMC 6900523. PMID 31740605.
- Shin, Seunggwan; Clarke, Dave J; Lemmon, Alan R; Moriarty Lemmon, Emily; Aitken, Alexander L; Haddad, Stephanie; Farrell, Brian D; Marvaldi, Adriana E; Oberprieler, Rolf G; McKenna, Duane D (2018-04-01). "Phylogenomic Data Yield New and Robust Insights into the Phylogeny and Evolution of Weevils". Molecular Biology and Evolution. 35 (4): 823–836. doi:10.1093/molbev/msx324. ISSN 0737-4038. PMID 29294021. S2CID 4366092.
- van Eldijk, Timo J. B.; Wappler, Torsten; Strother, Paul K.; van der Weijst, Carolien M. H.; Rajaei, Hossein; Visscher, Henk; van de Schootbrugge, Bas (January 2018). "A Triassic-Jurassic window into the evolution of Lepidoptera". Science Advances. 4 (1): e1701568. Bibcode:2018SciA....4.1568V. doi:10.1126/sciadv.1701568. ISSN 2375-2548. PMC 5770165. PMID 29349295.
- Huang, DiYing; Engel, Michael S.; Cai, ChenYang; Nel, André (May 2013). "Mesozoic giant fleas from northeastern China (Siphonaptera): Taxonomy and implications for palaeodiversity". Chinese Science Bulletin. 58 (14): 1682–1690. Bibcode:2013ChSBu..58.1682H. doi:10.1007/s11434-013-5769-3. ISSN 1001-6538. S2CID 53578959.
- Gao, Taiping; Shih, Chungkun; Rasnitsyn, Alexandr P.; Xu, Xing; Wang, Shuo; Ren, Dong (July 2013). "New Transitional Fleas from China Highlighting Diversity of Early Cretaceous Ectoparasitic Insects". Current Biology. 23 (13): 1261–1266. doi:10.1016/j.cub.2013.05.040. PMID 23810530. S2CID 9646168.
- Yang, Hongru; Shi, Chaofan; Engel, Michael S; Zhao, Zhipeng; Ren, Dong; Gao, Taiping (2020-04-02). "Early specializations for mimicry and defense in a Jurassic stick insect". National Science Review. 8: nwaa056. doi:10.1093/nsr/nwaa056. ISSN 2095-5138.
- Huang, Di-ying; Nel, André; Zompro, Oliver; Waller, Alain (2008-06-11). "Mantophasmatodea now in the Jurassic". Naturwissenschaften. 95 (10): 947–952. Bibcode:2008NW.....95..947H. doi:10.1007/s00114-008-0412-x. ISSN 0028-1042. PMID 18545982. S2CID 35408984.
- Selden, Paul A.; Baker, Anne S.; Phipps, Kenneth J. (2008). "An Oribatid Mite (arachnida: Acari) from the Oxford Clay (jurassic: Upper Callovian) of South Cave Station Quarry, Yorkshire, Uk". Palaeontology. 51 (3): 623–633. doi:10.1111/j.1475-4983.2008.00769.x. hdl:1808/8353. ISSN 1475-4983.
- Magalhaes, Ivan L. F.; Azevedo, Guilherme H. F.; Michalik, Peter; Ramírez, Martín J. (February 2020). "The fossil record of spiders revisited: implications for calibrating trees and evidence for a major faunal turnover since the Mesozoic". Biological Reviews. 95 (1): 184–217. doi:10.1111/brv.12559. ISSN 1464-7931. PMID 31713947. S2CID 207937170.
- Selden, Paul A.; Dunlop, Jason A. (2014). "The first fossil spider (Araneae: Palpimanoidea) from the Lower Jurassic (Grimmen, Germany)". Zootaxa. 3894 (1): 161–168. doi:10.11646/zootaxa.3894.1.13. PMID 25544628.
- Selden, P. A.; Shih, C.K.; Ren, D. (2013). "A giant spider from the Jurassic of China reveals greater diversity of the orbicularian stem group". Naturwissenschaften. 100 (12): 1171–1181. Bibcode:2013NW....100.1171S. doi:10.1007/s00114-013-1121-7. PMC 3889289. PMID 24317464.
- Dunlop, Jason A.; Kamenz, Carsten; Scholtz, Gerhard (June 2007). "Reinterpreting the morphology of the Jurassic scorpion Liassoscorpionides". Arthropod Structure & Development. 36 (2): 245–252. doi:10.1016/j.asd.2006.09.003. PMID 18089103.
- Huang, Diying; Selden, Paul A.; Dunlop, Jason A. (August 2009). "Harvestmen (Arachnida: Opiliones) from the Middle Jurassic of China". Naturwissenschaften. 96 (8): 955–962. Bibcode:2009NW.....96..955H. doi:10.1007/s00114-009-0556-3. ISSN 0028-1042. PMID 19495718. S2CID 9570512.
- Giribet, Gonzalo; Tourinho, Ana Lúcia; Shih, ChungKun; Ren, Dong (March 2012). "An exquisitely preserved harvestman (Arthropoda, Arachnida, Opiliones) from the Middle Jurassic of China". Organisms Diversity & Evolution. 12 (1): 51–56. doi:10.1007/s13127-011-0067-x. ISSN 1439-6092. S2CID 15658216.
- Dunhill, Alexander M.; Foster, William J.; Sciberras, James; Twitchett, Richard J. (January 2018). Hautmann, Michael (ed.). "Impact of the Late Triassic mass extinction on functional diversity and composition of marine ecosystems". Palaeontology. 61 (1): 133–148. doi:10.1111/pala.12332.
- Kiessling, Wolfgang (December 2009). "Geologic and Biologic Controls on the Evolution of Reefs". Annual Review of Ecology, Evolution, and Systematics. 40 (1): 173–192. doi:10.1146/annurev.ecolsys.110308.120251. ISSN 1543-592X.
- Klompmaker, A. A.; Schweitzer, C. E.; Feldmann, R. M.; Kowalewski, M. (2013-11-01). "The influence of reefs on the rise of Mesozoic marine crustaceans". Geology. 41 (11): 1179–1182. Bibcode:2013Geo....41.1179K. doi:10.1130/G34768.1. ISSN 0091-7613.
- Hudson, Wendy; Hart, Malcolm B.; Smart, Christopher W. (2009-01-01). "Palaeobiogeography of early planktonic foraminifera". Bulletin de la Société Géologique de France. 180 (1): 27–38. doi:10.2113/gssgfbull.180.1.27. ISSN 1777-5817.
- Wiggan, Nickolas J.; Riding, James B.; Fensome, Robert A.; Mattioli, Emanuela (2018-05-01). "The Bajocian (Middle Jurassic): A key interval in the early Mesozoic phytoplankton radiation". Earth-Science Reviews. 180: 126–146. Bibcode:2018ESRv..180..126W. doi:10.1016/j.earscirev.2018.03.009. ISSN 0012-8252.
- Zatoń, M.; Taylor, P.D. (2009-12-31). "Microconchids (Tentaculita) from the Middle Jurassic of Poland". Bulletin of Geosciences: 653–660. doi:10.3140/bull.geosci.1167. ISSN 1802-8225.
- Girard, Vincent; Saint Martin, Simona; Buffetaut, Eric; Saint Martin, Jean-Paul; Néraudeau, Didier; Peyrot, Daniel; Roghi, Guido; Ragazzi, Eugenio; Suteethorn, Varavudh (2020). Saint Martin, J.-P.; Saint Martin, S. (eds.). "Thai amber: insights into early diatom history?". BSGF - Earth Sciences Bulletin. 191: 23. doi:10.1051/bsgf/2020028. ISSN 1777-5817.
- Scholtz, Gerhard (November 2020). "Eocarcinus praecursor Withers, 1932 (Malacostraca, Decapoda, Meiura) is a stem group brachyuran". Arthropod Structure & Development. 59: 100991. doi:10.1016/j.asd.2020.100991. PMID 32891896.
- Schweitzer, Carrie E.; Feldmann, Rodney M. (2010-05-01). "The Oldest Brachyura (Decapoda: Homolodromioidea: Glaessneropsoidea) Known to Date (Jurassic)". Journal of Crustacean Biology. 30 (2): 251–256. doi:10.1651/09-3231.1. ISSN 0278-0372. S2CID 84707572.
- Guinot, Danièle (2019-11-14). "New hypotheses concerning the earliest brachyurans (Crustacea, Decapoda, Brachyura)". Geodiversitas. 41 (1): 747. doi:10.5252/geodiversitas2019v41a22. ISSN 1280-9659. S2CID 214220075.
- Fraaije, René; Schweigert, Günter; Nützel, Alexander; Havlik, Philipe (2013-01-01). "New Early Jurassic hermit crabs from Germany and France". Journal of Crustacean Biology. 33 (6): 802–817. doi:10.1163/1937240X-00002191. ISSN 0278-0372.
- Mironenko, Aleksandr (January 2020). "A hermit crab preserved inside an ammonite shell from the Upper Jurassic of central Russia: Implications to ammonoid palaeoecology". Palaeogeography, Palaeoclimatology, Palaeoecology. 537: 109397. Bibcode:2020PPP...537j9397M. doi:10.1016/j.palaeo.2019.109397.
- Bracken-Grissom, Heather D.; Ahyong, Shane T.; Wilkinson, Richard D.; Feldmann, Rodney M.; Schweitzer, Carrie E.; Breinholt, Jesse W.; Bendall, Matthew; Palero, Ferran; Chan, Tin-Yam; Felder, Darryl L.; Robles, Rafael (2014-07-01). "The Emergence of Lobsters: Phylogenetic Relationships, Morphological Evolution and Divergence Time Comparisons of an Ancient Group (Decapoda: Achelata, Astacidea, Glypheidea, Polychelida)". Systematic Biology. 63 (4): 457–479. doi:10.1093/sysbio/syu008. ISSN 1063-5157. PMID 24562813.
- Braig, Florian; Haug, Joachim T.; Schädel, Mario; Haug, Carolin (2019-06-28). "A new thylacocephalan crustacean from the Upper Jurassic lithographic limestones of southern Germany and the diversity of Thylacocephala". Palaeodiversity. 12 (1): 69. doi:10.18476/pale.v12.a6. ISSN 1867-6294.
- Vörös, Attila; Kocsis, Ádám T.; Pálfy, József (2019). "Mass extinctions and clade extinctions in the history of brachiopods: Brief review and a post-Paleozoic case study". Rivista Italiana di Paleontologia e Stratigrafia. 125 (3). doi:10.13130/2039-4942/12184. ISSN 2039-4942.
- Manojlovic, Marko; Clapham, Matthew E. (2020-11-23). "The role of bioturbation-driven substrate disturbance in the Mesozoic brachiopod decline". Paleobiology: 1–15. doi:10.1017/pab.2020.50. ISSN 0094-8373.
- Page, Kevin N. (January 2008). "The evolution and geography of Jurassic ammonoids". Proceedings of the Geologists' Association. 119 (1): 35–57. doi:10.1016/S0016-7878(08)80257-X.
- Ros, Sonia; De Renzi, Miquel; Damborenea, Susana E.; Márquez-Aliaga, Ana (November 2011). "Coping between crises: Early Triassic–early Jurassic bivalve diversity dynamics". Palaeogeography, Palaeoclimatology, Palaeoecology. 311 (3–4): 184–199. Bibcode:2011PPP...311..184R. doi:10.1016/j.palaeo.2011.08.020.
- Sha, J.; Cestari, R.; Fabbi, S. (April 2020). "Paleobiogeographic distribution of rudist bivalves (Hippuritida) in the Oxfordian–early Aptian (Late Jurassic–Early Cretaceous)". Cretaceous Research. 108: 104289. doi:10.1016/j.cretres.2019.104289.
- Iba, Yasuhiro; Sano, Shin-ichi; Mutterlose, Jörg (2014-05-02). Samonds, Karen E. (ed.). "The Early Evolutionary History of Belemnites: New Data from Japan". PLOS ONE. 9 (5): e95632. Bibcode:2014PLoSO...995632I. doi:10.1371/journal.pone.0095632. ISSN 1932-6203. PMC 4008418. PMID 24788872.
- Hoffmann, René; Stevens, Kevin (February 2020). "The palaeobiology of belemnites – foundation for the interpretation of rostrum geochemistry". Biological Reviews. 95 (1): 94–123. doi:10.1111/brv.12557. ISSN 1464-7931. PMID 31729839. S2CID 208036104.
Sources
- Arkell, W.J. (1956). Jurassic Geology of the World. Edinburgh: Oliver & Boyd.
- Behrensmeyer, A.K.; Damuth, J.D.; DiMichele, W.A.; Potts, R.; Sues, H.D.; Wing, S.L., eds. (1992). Terrestrial Ecosystems through Time: the Evolutionary Paleoecology of Terrestrial Plants and Animals. Chicago: University of Chicago Press. ISBN 0-226-04154-9.
- Brongniart, Alexandre (1829). Tableau des terrains qui composent l'écorce du globe ou essai sur la structure de la partie connue de la terre [Description of the Terrains that Constitute the Crust of the Earth or Essay on the Structure of the Known Lands of the Earth] (in French). Strasbourg – via Gallica.
- Carroll, R. L. (1988). Vertebrate Paleontology and Evolution. New York: WH Freeman. ISBN 978-0-7167-1822-2.
- Feduccia, A. (1996). The Origin and Evolution of Birds. New Haven: Yale University Press. ISBN 978-0-300-06460-5.
- Gannon, Megan (31 October 2012). "Jurassic turtle graveyard found in China". cbsnews.com. Retrieved 10 October 2019.
- Haines, Tim (2000). Walking with Dinosaurs: A Natural History. New York: Dorling Kindersley. ISBN 978-0-7894-5187-3.
- Hallam, A. (1986). "The Pliensbachian and Tithonian extinction events". Nature. 319 (6056): 765–768. Bibcode:1986Natur.319..765H. doi:10.1038/319765a0. S2CID 4310433.
- Hölder, H. (1964). Jura – Handbuch der stratigraphischen Geologie (in German). IV. Stuttgart: Enke-Verlag.
- Jacobs, Louis, L. (1997). "African Dinosaurs". In Currie, Phillip J.; Padian, Kevin (eds.). Encyclopedia of Dinosaurs. Academic Press.
- Jaramillo, Jessica (March–April 2014). "Entrevista al Dr. Víctor Alberto Ramos, Premio México Ciencia y Tecnología 2013" (in Spanish). Vol. 17 no. 66.
Si logramos publicar esos nuevos resultados, sería el primer paso para cambiar formalmente la edad del Jurásico-Cretácico. A partir de ahí, la Unión Internacional de la Ciencias Geológicas y la Comisión Internacional de Estratigrafía certificaría o no, depende de los resultados, ese cambio.
Cite magazine requires|magazine=(help) - Kazlev, M. Alan (2002). "Palaeos Mesozoic: Jurassic: The Jurassic Period". Palaeos. Archived from the original on 5 January 2006. Retrieved January 8, 2006.
- Pieńkowski, G.; Schudack, M.E.; Bosák, P.; Enay, R.; Feldman-Olszewska, A.; Golonka, J.; Gutowski, J.; et al. (2008). "Jurassic". In McCann, T. (ed.). The Geology of Central Europe. Mesozoic and Cenozoic. London: Geological Society.
- Rollier, L. (1903). "Das Schweizerische Juragebirge". Sonderabdruck aus dem Geographischen Lexikon der Schweiz (in German). Attinger, Neuenburg: von Gebr.
- Monroe, James S.; Wicander, Reed (1997). The Changing Earth: Exploring Geology and Evolution (2nd ed.). Belmont: West Publishing Company. ISBN 0-314-09577-2.
- Motani, R. (2000). "Rulers of the Jurassic Seas". Scientific American. Vol. 283 no. 6. doi:10.1038/scientificamerican1200-52.
- Scotese, Christopher R. (2003). "Pangea Begins to Rift Apart". scotese.com. Retrieved 2019-10-10.
- Stanley, S.M.; Hardie, L.A. (1998). "Secular oscillations in the carbonate mineralogy of reef-building and sediment-producing organisms driven by tectonically forced shifts in seawater chemistry". Palaeogeography, Palaeoclimatology, Palaeoecology. 144 (1–2): 3–19. Bibcode:1998PPP...144....3S. doi:10.1016/s0031-0182(98)00109-6.
- Taylor, P.D.; Wilson, M.A. (2003). "Palaeoecology and evolution of marine hard substrate communities" (PDF). Earth-Science Reviews. 62 (1): 1–103. Bibcode:2003ESRv...62....1T. doi:10.1016/s0012-8252(02)00131-9. Archived from the original (PDF) on 2009-03-25.
- Vennari, Verónica V.; Lescano, Marina; Naipauer, Maximiliano; Aguirre-Urreta, Beatriz; Concheyro, Andrea; Schaltegger, Urs; Armstrong, Richard; Pimentel, Marcio; Ramos, Victor A. (2014). "New constraints on the Jurassic–Cretaceous boundary in the High Andes using high-precision U–Pb data". Gondwana Research. 26 (1): 374–385. Bibcode:2014GondR..26..374V. doi:10.1016/j.gr.2013.07.005.
- Vinn, O.; ten Hove, H.A.; Mutvei, H. (2008). "On the tube ultrastructure and origin of calcification in sabellids (Annelida, Polychaeta)". Palaeontology. 51 (2): 295–301. doi:10.1111/j.1475-4983.2008.00763.x.
- Vinn, O.; Mutvei, H. (2009). "Calcareous tubeworms of the Phanerozoic" (PDF). Estonian Journal of Earth Sciences. 58 (4): 286–296. doi:10.3176/earth.2009.4.07. Gale A216178929.
- von Humboldt, Alexander (1799). Ueber die unterirdischen Gasarten und die Mittel, ihren Nachteil zu vermindern, ein Beitrag zur Physik der praktischen Bergbaukunde [On the types of subterranean gases and means of minimizing their harm, a contribution to the physics of practical mining] (in German). Braunschweig: Vieweg.
- von Humboldt, Alexander (1858). Kosmos (in German). 4. Stuttgart: Cotta. p. 632 – via HathiTrust.
- Wings, Oliver; Rabi, Márton; Schneider, Jörg W.; Schwermann, Leonie; Sun, Ge; Zhou, Chang-Fu; Joyce, Walter G. (2012). "An enormous Jurassic turtle bone bed from the Turpan Basin of Xinjiang, China". Naturwissenschaften. 114 (11): 925–935. Bibcode:2012NW.....99..925W. doi:10.1007/s00114-012-0974-5. PMID 23086389. S2CID 17423081.
- Witton, Mark P.; Martill, David M.; Loveridge, Robert F. (2010). "Clipping the Wings of Giant Pterosaurs: Comments on Wingspan Estimations and Diversity". Acta Geoscientica Sinica. 31 (Supp 1): 79–81.
- Witton, Mark P. (23 June 2016). "Why the giant azhdarchid Arambourgiania philadelphiae needs a fanclub". markwitton-com.blogspot.com.
Further reading
- Mader, Sylvia (2004). Biology (eighth ed.).
- Ogg, Jim (June 2004). Overview of Global Boundary Stratotype Sections and Points (GSSP's). International Commission on Stratigraphy. p. 17.
- Stanley, S.M.; Hardie, L.A. (1999). "Hypercalcification; paleontology links plate tectonics and geochemistry to sedimentology". GSA Today. 9: 1–7.
External links
| Wikimedia Commons has media related to Jurassic. |
| Look up jurassic in Wiktionary, the free dictionary. |
- Examples of Jurassic Fossils
- Jurassic (chronostratigraphy scale)
- Jurassic fossils in Harbury, Warwickshire
- Jurassic Microfossils: 65+ images of Foraminifera
- . Encyclopædia Britannica. 15 (11th ed.). 1911. With map and table.