Basilosaurus
Basilosaurus (meaning "king lizard") is a genus of large, predatory, prehistoric archaeocete whale from the late Eocene, approximately 41.3 to 33.9 million years ago (mya). First described in 1834, it was the first archaeocete and prehistoric whale known to science.[2] The first fossils were discovered along the Gulf Coast of the United States, along with a few fossils in the eastern U.S., attributed to the type species B. cetoides. They were originally thought to be of a giant reptile, hence the suffix "-saurus", Ancient Greek for "lizard". The animal was later found to be an early marine mammal, which prompted attempts at renaming the creature, which failed as zoological nomenclature dictates using the original name given. Fossils were later found of the second species, B. isis, in 1904 in North Africa, more specifically the countries of Egypt, Jordan, Tunisia[3] and Morocco. Basilosaurus has a possible record in Seymour Island in Antarctica.[4]
| Basilosaurus | |
|---|---|
.jpg.webp) | |
| B. cetoides skeleton, National Museum of Natural History, Washington DC | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Artiodactyla |
| Infraorder: | Cetacea |
| Family: | †Basilosauridae |
| Subfamily: | †Basilosaurinae |
| Genus: | †Basilosaurus Harlan 1834 |
| Species | |
| |
| Synonyms | |
Basilosaurus is considered to have been common in the Tethys sea.[5][6] It was one of the largest, if not the largest, animals of the Paleogene. It was the top predator of its environment, preying on sharks, large fish and other marine mammals, namely the dolphin-like Dorudon, which seems to have been their predominant food source.
Basilosaurus was at one point a wastebasket taxon, before the genus slowly started getting reevaluated, with many species of different Eocene cetacean being assigned to the genus in the past, however they are invalid or have been reclassified under a new or different genus, leaving only 2 confirmed species. Basilosaurus may have been one of the first fully aquatic cetaceans[2] (sometimes referred to as the pelagiceti[7]). Basilosaurus, unlike modern cetaceans, had various types of teeth–such as canines and molars–in its mouth (heterodonty), and it probably was able to chew its food in contrast to modern cetaceans which swallow their food whole.[8][9]
Description

Measuring 15–20 m (49–66 ft),[10][11][12][13] Basilosaurus is one of the largest-known animals to exist between the K–Pg extinction event 66 million years ago (mya) and around 15 million years ago when modern cetaceans began to reach enormous sizes.[14] B. isis is slightly smaller than B. cetoides,[15] with B. isis being 15–18 m (49–59 ft) long and B. cetoides being 17–20 m (56–66 ft) long.[12][13] Basilosaurus is distinguished from other basilosaurids genera by its larger body size and its more elongated posterior thoracic, lumbar, and anterior caudal vertebrae. Basilosaurus does not have the vertically oriented metapophyses seen in its closest relative the basilosaurid known as Basiloterus. Basilosaurus is considered to be the largest of archeocete whales.[16][17]
Cranium

The dental formula for B. isis is 3.1.4.23.1.4.3. The upper and lower molars and second to fourth premolars are double-rooted and high-crowned.[18]
The head of Basilosaurus did not have room for a melon like modern toothed whales, and the brain was smaller in comparison, as well. They are not believed to have had the social capabilities of modern whales.
A 2011 study concluded that the skull of Basilosaurus is asymmetrical like in modern toothed whales, and not, as previously assumed, symmetrical like in baleen whales and artiodactyls (which are closely related to cetaceans). In modern toothed whales, this asymmetry is associated with high-frequency sound production and echolocation, neither of which is thought to have been present in Basilosaurus. This probably evolved to detect sound underwater, with a fatty sound-receiving pad in the mandible.[19]
In the skull, the inner and middle ear are enclosed by a dense tympanic bulla.[20] The synapomorphic cetacean air sinus system is partially present in basilosaurids, including the pterygoid, peribullary, maxillary, and frontal sinuses.[21] The periotic bone, which surrounds the inner ear, is partially isolated. The mandibular canal is large and laterally flanked by a thin bony wall, the pan bone or acoustic fenestra. These features enabled basilosaurs to hear directionally in water.[20]
The ear of basilosaurids is more derived than those in earlier archaeocetes, such as remingtonocetids and protocetids, in the acoustic isolation provided by the air-filled sinuses inserted between the ear and the skull. The basilosaurid ear did, however, have a large external auditory meatus, strongly reduced in modern cetaceans, but, though this was probably functional, it can have been of little use under water.[22]
Hind limbs

A 16-meter (52 ft) individual of B. isis had 35-centimeter-long (14 in) hind limbs with fused tarsals and only three digits. The limited size of the limb and the absence of an articulation with the sacral vertebrae make a locomotory function unlikely.[23] Analysis has shown that the reduced limbs could rapidly adduct between only two positions.[24] Possible uses for the structure have been given, such as clasper-like body functions (compare to the function of pelvic spurs, the last vestiges of limbs in certain modern snakes). These limbs would have been used to guide the animals' long bodies during mating.[25]
Spine and movement
A complete Basilosaurus skeleton was found in 2015, and several attempts have been made to reconstruct the vertebral column from partial skeletons. Kellogg 1936 estimated a total of 58 vertebrae, based on two partial and nonoverlapping skeletons of B. cetoides from Alabama. More complete fossils uncovered in Egypt in the 1990s allowed a more accurate estimation: the vertebral column of B. isis has been reconstructed from three overlapping skeletons to a total of 70 vertebrae with a vertebral formula interpreted as seven cervical, 18 thoracic, 20 lumbar and sacral, and 25 caudal vertebrae. The vertebral formula of B. cetoides can be assumed to be the same.[26]

Basilosaurus has an anguilliform (eel-like) body shape because of the elongation of the centra of the thoracic through anterior caudal vertebrae. In life, these vertebrae were filled with marrow, and because of the enlarged size, this made them buoyant. Basilosaurus probably swam predominantly in two dimensions at the sea surface, in contrast to the smaller Dorudon, which was likely a diving, three-dimensional swimmer.[27] The skeletal anatomy of the tail suggests that a small fluke was probably present, which would have aided only vertical motion.[28]
Similarly sized thoracic, lumbar, sacral, and caudal vertebrae imply that it moved in an anguilliform fashion, but predominantly in the vertical plane. Paleontologist Philip D. Gingerich theorized that Basilosaurus may also have moved in a very odd, horizontal anguilliform fashion to some degree, something completely unknown in modern cetaceans. The vertebrae appear to have been hollow, and likely also fluid-filled. This would imply that Basilosaurus typically functioned in only two dimensions at the ocean surface, compared with the three-dimensional habits of most other cetaceans. Judging from the relatively weak axial musculature and the thick bones in the limbs, Basilosaurus is not believed to have been capable of sustained swimming or deep diving, or terrestrial locomotion.[29] Basilosaurus did still have an elbow joint in its flipper like a seal.
Paleobiology
Feeding
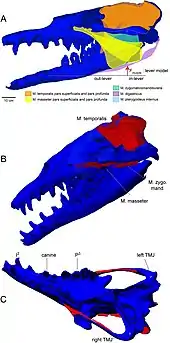
The cheek teeth of Basilosaurus retain a complex morphology and functional occlusion. Heavy wear on the teeth reveals that food was first chewed then swallowed.[20] Scientists were able to estimate the bite force of Basilosaurus by analyzing the scarred skull bones of another species of prehistoric whale, Dorudon, and concluded they could bite with a force of 3,600 pounds per square inch (25 MPa).[30]
Analyses of the stomach contents of B. cetoides has shown that this species fed exclusively on fish and large sharks, while bite marks on the skulls of juvenile Dorudon have been matched with the dentition of B. isis, suggesting a dietary difference between the two species, similar to that found in different populations of modern killer whales.[18] It was probably an active predator rather than a scavenger.[31] The discovery of juvenile Dorudon at Wadi Al Hitan bearing distinctive bite marks on their skulls indicates that B. isis would have aimed for the skulls of its victims to kill its prey, and then subsequently torn its meals apart, based on the disarticulated remains of the Dorudon skeletons. The finding further cements theories that B. isis was an apex predator that may have hunted newborn and juvenile Dorudon at Wadi Al Hitan when mothers of the latter came to give birth.[12] The stomach contents of an elderly male B. isis not only includes Dorudon but the fish Pycnodus mokattamensis.[12]
Paleoecology
Basilosaurus would have been the top predator of its environment.[32] Basilosaurus lived in the warm tropical environment of the Eocene in areas abundant with sea grasses, such as Thalassodendron, Thalassia (also known as turtle grass) and Halodule.[33][34][35] Basilosaurus would have coexisted with the dolphin-like Dorudon,[36] the whales Cynthiacetus and Basiloterus,[37] the primitive sirenian Protosiren, the early elephant Moeritherium,[38][39][40] the sea turtle Puppigerus[41] and many sharks, such as Galeocerdo alabamensis,[42] Physogaleus, Otodus, Squatina prima, Striatolamia, Carcharocles sokolovi and Isurus praecursor.[43]
Extinction
Basilosaurus fossil record seems to end at about 35–33.9 mya.[44] Basilosaurus extinction coincides with the Eocene–Oligocene extinction event which happened 33.9 mya,[45] which also resulted in the extinction of almost all other archaeocetes.[13] The event has been attributed to volcanic activity, meteor impacts or a sudden change in climate (such as the environment getting cooler), the latter of which might have caused changes in the ocean by disrupting oceanic circulation.[15][46][47] Basilosaurus went extinct leaving no relatives. Despite this new currents and deep ocean upwelling favored the diversification of modern cetaceans such as early toothed and baleen whales.
Classification
Below is the phylogenetic analysis on the placement of Basilosaurus. Two subfamilies exist in Basilosauridae: Basilosaurinae which includes Basilosaurus, and Dorudontinae. These groups have been declared invalid in the past.[48][49] Dorudon remains were once thought to represent juvenile Basilosaurus.[50]
| †Basilosaurinae |
| |||||||||||||||||||||
Taxonomic history
Etymology

The two species of Basilosaurus are B. cetoides, whose remains were discovered in the United States, and B. isis, which was discovered in Egypt. B. cetoides is the type species for the genus.[26][51] The Holotype Harlan found, was found in Ouachita County, Louisiana.[52][53] Vertebrae were sent to the American Philosophical Society by a Judge Henry Bry of Ouachita County, Louisiana and Judge John Creagh of Clarke County, Alabama. Both fossils ended up in the hands of the anatomist Richard Harlan, who requested more examples from Creagh.[54][55] The first bones were unearthed when rain caused a hillside full of sea shells to slide. The bones were lying in a curved line "measuring upwards of four hundred feet in length, with intervals which were vacant." Many of these bones were used as andirons and destroyed; Bry saved the bones he could find, but was convinced more bones were still to be found on the location. Bry speculated that the bones must have belonged to a "sea monster" and supplied "a piece having the appearance of a tooth" to help determine which kind.[56] Harlan identified the tooth as a wedge-shaped shell and instead focused on "a vertebra of enormous dimensions" which he assumed belonged to the order "Enalio-Sauri of Conybeare", "found only in the sub-cretaceous series."[57] He noted that some parts of the vertebra were similar to those of Plesiosaurus and skull was similar to Mosasaurus, but that they were completely different in proportions. Comparing his vertebra to those of large dinosaurs such as Megalosaurus and Iguanodon, Harlan concluded that his specimen was considerably larger—he estimated the animal to have been no less than 80–100 ft (24–30 m) long—and therefore suggested the name Basilosaurus, meaning "king lizard".[58]
Harlan brought his assembled specimens (including fragments of jaw and teeth, humerus, and rib fragments) to the UK where he presented them to anatomist Richard Owen. Owen concluded that the molar teeth were two-rooted, a dental morphology unknown in fishes and reptiles, and more complex and varied than in any known reptile, and therefore that the specimen must be a mammal. Owen correctly associated the teeth with cetaceans, but he thought it was an herbivorous animal, similar to sirenians.[59] Consequently, Owen proposed renaming the find Zeuglodon cetoides ("whale-like yoke teeth" in reference to the double-rooted teeth) and Harlan agreed.[60]
Wadi El Hitan


Wadi El Hitan, Arabic وادي الحيتان , "Valley of the Whales", is an Egyptian sandstone formation where many early-whale skeletons were discovered.[61] German botanist Georg August Schweinfurth discovered the first archaeocete whale in Egypt (Zeuglodon osiris, now Saghacetus osiris) in 1879. He visited the Qasr el Sagha Formation in 1884 and 1886 and missed the now famous Wadi El Hitan by a few kilometers. German paleontologist Wilhelm Barnim Dames described the material, including the type specimen of Z. osiris, a well-preserved dentary.[11]
Hugh Beadnell, head of the Geological Survey of Egypt 1896–1906,[11] named and described Zeuglodon isis in Andrews 1904 based on a partial mandible and several vertebrae from Wadi El Hitan in Egypt.[62] Andrews 1906[63] described a skull and some vertebrae of a smaller archaeocete and named it Prozeuglodon atrox, now known today as Dorudon atrox. Kellogg 1936 discovered deciduous teeth in this skull and it was then believed to be a juvenile [Pro]zeuglodon isis for decades before more complete fossils of mature Dorudon were discovered.[64][11][65]
In the 1980s, Elwyn L. Simons and Philip D. Gingerich started to excavate at Qasr el-Sagha and Wadi El Hitan with the hope of finding material that could match archaeocete fossils from Pakistan. Since then, over 500 archaeocete skeletons have been found at these two locations, of which most are B. isis or D. atrox, several of the latter carrying bite marks assumed to be from the former.[11] A 1990 paper described additional fossils including foot bones and speculated that the reduced hind limbs were used as copulatory guides.[24] One thing that was noted, was that whale fossils were so common, that when a mason company looked at their newest table counter, they realized that they had created a cross section of a 40 million year old Basilosaurid fossil. This find was another thing that caught the eye of Gingerich.[66]
In 2016, a complete skeleton, the first-ever find for Basilosaurus, was uncovered in Wadi El Hitan, preserved with the remains of its prey, including a Dorudon and several species of fish.[12] The whale's skeleton also shows signs of scavenging or predation by large sharks such as the otodontid Carcharocles sokolovi.
Wastebasket taxa
Many dubious species have been assigned to Basilosaurus in the past which have since been invalidated or were too incomplete to determine anything.
Nomina dubia
A nomen dubium is a scientific name that is of unknown or doubtful application. There are a few documented cases of this being applied to Basilosaurus in the past.

- Zeuglodon wanklyni, was a supposed species of Basilosaurus, that described in 1876 based on a skull found in the Wanklyn's Barton Cliff in the United Kingdom. This single specimen, however, quickly disappeared and has since been declared a nomen nudum or referred to as Zygorhiza wanklyni.[67]
- Zeuglodon vredense or vredensis was named in the 19th century based on a single, isolated tooth without any kind of accompanying description, and Kellogg 1936 therefore declared it a nomen nudum.[68]</ref>[69]
- Zeuglodon puschi[i] was a species that was said to come from Poland, it was named by Brandt 1873. Kellogg 1936 noted that the species is based on an incomplete vertebra of indeterminable position and, therefore, that the species is invalid.[70][71]
- Zeuglodon brachyspondylus was described by Johannes Peter Müller based on some vertebrae from "Zeuglodon hydrarchus",[72] better known as Dr. Albert Koch's "Hydrarchos". Kellogg 1936, synonymized it with Pontogeneus priscus, which a 2005 study declared a nomen dubium.[73]
Reassigned species

- Basilosaurus drazindai was named by a 1997 study based on a single lumbar vertebra. Originally, the species was thought to have lived in Pakistan and the UK.[16] It was later declared a nomen dubium by Uhen (2013), but Gingerich and Zouhri (in press) reassigned it to the genus Eocetus. This species was at one point in time concluded to be the earliest record of the genus Basilosaurus, before its reclassification.[74][75][76]
- Zeuglodon elliotsmithii, Z. sensitivius, Z. sensitivus, and Z. zitteli were synonymized and grouped under the genus Saghacetus by a 1992 study.[77]
- Zeuglodon paulsoni from Ukraine (then the Russian Empire) was named by Brandt 1873. It was synonymized with Platyosphys but is now considered nomen dubium. Gingerich and Zouhri (in press), however, maintain Platyosphys as valid.[76][78]
- Basilosaurus caucasicus also known as Basilosaurus caucasicum or Zeuglodon caucasicum was species described in the Russian Empire, it gets its name from the Russian caucasus of where it was found in the 1890s.[79] The specimen was found. The fossil was reassigned to the toothed whale Microzeuglodon caucasicum.[80]
- Basilosaurus harwoodi was discovered in Murray River near Wellington in South Australia, this species classification is quite controversial, yet today it is agreed that the Australian, Basilosaurus harwoodi (or Zeuglodon harwoodi) is most likely a member of the genus Metasqualodon as noted by T. S. Hall 1911.[81][82]
- In 1906, German naturalist Othenio Abel thought fossils from the Eocene of Alabama, previously described in 1900 as being a Basilosaurus hip bone by American zoologist Frederic Augustus Lucas, represented the shoulder of a large bird similar to Gastornis, and named it Alabamornis gigantea.[83] Lucas later countered his conclusion in 1908 as he reassigned the fossil specimens to the original conclusion of a Basilosaurus hip bone.[84]
In popular culture
The species B. cetoides is the state fossil of Alabama[15] and Mississippi.[85][86] Basilosaurus is featured in the BBC's Walking with series in Walking with Beasts and Sea Monsters.[87][88][89] During the early 19th century, B. cetoides fossils were so common (and sufficiently large) that they were regularly used as furniture in the American South.[90]
See also
References
- "Dir Abu Lifa (Eocene of Egypt)". PBDB. Mark Uhen.
- Uhen, Mark D. (2002). "Basilosaurids". In Perrin, William F.; Würsig, Bernd; Thewissen, J. G. M. (eds.). Encyclopedia of Marine Mammals. Academic Press. pp. 79–81. ISBN 978-0-12-551340-1.
- Werdelin, Lars; Sanders, William Joseph (2010-07-20). Cenozoic Mammals of Africa. ISBN 9780520257214.
- "Antarctic Paleontological Heritage: Late Cretaceous–Paleogene vertebrates from Seymour (Marambio) Island, Antarctic Peninsula". Advances in Polar Science. 30. 2019-08-20. doi:10.13679/j.advps.2019.0015.
- "Valley of the Whales (magazine)". National Geographic. August 2010.
- "Basilosaurus FOSSIL MAMMAL GENUS". Encyclopedia Britannica.
- "Clade Pelagiceti Uhen 2008 (whale)". PBDB.
- "HOW DOES DIGESTION TAKE PLACE IN WHALES?". Whales, online. 2015-07-15.
- "Basilosaurus the Bone-Crusher". National geographic. 2015-03-06.
- Gingerich, P. D. (2012). "Evolution of Whales from Land to Sea" (PDF). Proceedings of the American Philosophical Society. 156 (3): 309–323.
- Gingerich, P. D. (2008). "Early Evolution of Whales: A Century of Research in Egypt" (PDF). In Fleagle, J. G.; Gilbert, C. C. (eds.). Elwyn Simons: A Search for Origins. Developments in Primatology: Progress and Prospects. Berlin: Springer. pp. 107–124. ISBN 978-0-387-73895-6.
- Voss, Manja; Antar, Mohammed Sameh M.; Zalmout, Iyad S.; Gingerich, Philip D. (2019). "Stomach contents of the archaeocete Basilosaurus isis: Apex predator in oceans of the late Eocene". PLOS ONE. 14 (1). e0209021. Bibcode:2019PLoSO..1409021V. doi:10.1371/journal.pone.0209021. PMC 6326415. PMID 30625131.
- Kellogg 1936, p. .
- "Explore Our Collections Basilosaurus". Smithsonian natural history museum. Archived from the original on 2016-03-31.
- Gingerich, Philip D. "Basilosaurus cetoides". Encyclopedia of Alabama. Retrieved 24 August 2013.
- Gingerich, P. D.; Arif, M; Bhatti, M Akram; Anwar, M; Sanders, William J (1997). "Basilosaurus drazindai and Basiloterus hussaini, New Archaeoceti (Mammalia, Cetacea) from the Middle Eocene Drazinda Formation, with a Revised Interpretation of Ages of Whale-Bearing Strata in the Kirthar Group of the Sulaiman Range, Punjab (Pakistan)". Contributions from the Museum of Paleontology, University of Michigan. 30 (2): 55–81. hdl:2027.42/48652. OCLC 742731913.
- "Basilosaurus description". PBDB.
- Fahlke, Julia M. (2012). "Bite marks revisited – evidence for middle-to-late Eocene Basilosaurus isis predation on Dorudon atrox (both Cetacea, Basilosauridae)" (PDF). Palaeontologia Electronica. 15 (3).
- Fahlke, Julia M.; Gingerich, Philip D.; Welsh, Robert C.; Wood, Aaron R. (2011). "Cranial asymmetry in Eocene archaeocete whales and the evolution of directional hearing in water". PNAS. 108 (35): 14545–14548. Bibcode:2011PNAS..10814545F. doi:10.1073/pnas.1108927108. PMC 3167538. PMID 21873217.
- Gingerich, P. D.; Uhen, Mark D. (1998). "Likelihood estimation of the time of origin of Cetacea and the time of divergence of Cetacea and Artiodactyla" (PDF). Palaeontologia Electronica. 1 (2): 4.
- Racicot, Rachel A.; Berta, Annalisa (2013). "Comparative Morphology of Porpoise (Cetacea: Phocoenidae) Pterygoid Sinuses: Phylogenetic and Functional Implications". Journal of Morphology. 274 (1): 50. doi:10.1002/jmor.20075. PMID 22965565. S2CID 2617769.
- Nummela, Sirpa; Thewissen, J. G. M.; Bajpai, Sunil; Hussain, Taseer; Kumar, Kishor (2004). "Eocene evolution of whale hearing". Nature. 430 (7001): 776–778. Bibcode:2004Natur.430..776N. doi:10.1038/nature02720. PMID 15306808. S2CID 4372872.
- Bejder, Lars; Hall, Brian K. (2002). "Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss" (PDF). Evolution and Development. 4 (6): 445–458. doi:10.1046/j.1525-142x.2002.02033.x. PMID 12492145. S2CID 8448387.
- Gingerich, Philip D.; Smith, B. Holly; Simons, Elwyn L. (1990). "Hind limbs of eocene basilosaurus: evidence of feet in whales". Science. 249 (4965): 154–157. Bibcode:1990Sci...249..154G. doi:10.1126/science.249.4965.154. PMID 17836967. S2CID 35307146.
- "Basilosaurid FOSSIL MAMMAL". Encyclopedia Britannica.
- Zalmout, I. S.; Mustafa, H. A.; Gingerich, P. D. (2000). "Priabonian Basilosaurus isis (Cetacea) from the Wadi Esh-Shallala Formation: first marine mammal from the Eocene of Jordan". Journal of Vertebrate Paleontology. 20 (1): 201–204. doi:10.1671/0272-4634(2000)020[0201:pbicft]2.0.co;2. OCLC 4908948040.
- Gingerich, P. D. (1998). "Paleobiological Perspectives on Mesonychia, Archaeoceti, and the Origin of Whales" (PDF). In Thewissen, J. G. M. (ed.). The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea. Advances in Vertebrate Paleobiology. 1. Springer. pp. 424–439. ISBN 9780306458538.
- "Basilosaurus spp". College of Osteopathic Medicine. Robert Boessenecker and Jonathan Geisler.
- Great Transformations in Vertebrate Evolution. KENNETH P. DIAL, NEIL SHUBIN, AND ELIZABETH L. BRAINERD. July 2015. ISBN 9780226268255.
- Snively, E.; Fahlke, J. M.; Welsh, R. C. (2015). "Bone-Breaking Bite Force of Basilosaurus isis (Mammalia, Cetacea) from the Late Eocene of Egypt Estimated by Finite Element Analysis". PLOS ONE. 10 (2): e0118380. Bibcode:2015PLoSO..1018380S. doi:10.1371/journal.pone.0118380. PMC 4340796. PMID 25714832.
- Snively, Eric; Fahlke, Julia M.; Welsh, Robert C. (25 February 2015). "Bone-Breaking Bite Force of Basilosaurus isis (Mammalia, Cetacea) from the Late Eocene of Egypt Estimated by Finite Element Analysis". PLOS ONE. 10 (2): e0118380. Bibcode:2015PLoSO..1018380S. doi:10.1371/journal.pone.0118380. PMC 4340796. PMID 25714832.
- "15-meter-long ancient whale Basilosaurus isis was top marine predator". Science Daily.
- "Florida Lime Works quarry (Eocene of the United States)". PBDB.
- "Wadi Hitan ZV-74 (Eocene of Egypt)". PBDB.
- "Eocene Epoch". Encyclopedia Britannica.
- "†Dorudon Gibbes 1845 (whale)". PBDB.
- "†Cynthiacetus Uhen 2005 (whale)". PBDB.
- The topography and geology of the Fayum province of Egypt. Cairo: National Printing Dept. 1905. pp. 87–88.
- Holroyd, Patricia A.; Parham, James F.; Hutchison, J. Howard (2005). "A Reappraisal of Some Paleogene Turtles from the Southeastern United States". Journal of Vertebrate Paleontology. 25 (4): 979–982. doi:10.1671/0272-4634(2005)025[0979:AROSPT]2.0.CO;2. JSTOR 4524524.
- "North American Eocene Sea Cows (Mammalia: Sirenia)". Daryl P.; Morgan, Gary S.; Ray, Clayton E. hdl:10088/1974. Cite journal requires
|journal=(help) - "†Puppigerus Cope 1870 (sea turtle)". PBDB.
- "†Galeocerdo alabamensis Leriche 1942 (tiger shark)". Fossilworks.
- "Hardie Kaolin Mine (Eocene of the United States)". PBDB.
- "†Basilosaurus Harlan 1834 (whale)". PBDB.
- Ivany, Linda C.; Patterson, William P.; Lohmann, Kyger C. (2000). "Cooler winters as a possible cause of mass extinctions at the Eocene/Oligocene boundary" (PDF). Nature. 407 (6806): 887–890. Bibcode:2000Natur.407..887I. doi:10.1038/35038044. hdl:2027.42/62707. PMID 11057663. S2CID 4408282.
- "Russia's Popigai Meteor Crash Linked to Mass Extinction". Yahoo news. Becky Oskin, Senior Writer.
- Molina, Eustoquio; Gonzalvo, Concepción; Ortiz, Silvia; Cruz, Luis E. (2006-02-28). "Foraminiferal turnover across the Eocene–Oligocene transition at Fuente Caldera, southern Spain: No cause–effect relationship between meteorite impacts and extinctions". Marine Micropaleontology. 58 (4): 270–286. Bibcode:2006MarMP..58..270M. doi:10.1016/j.marmicro.2005.11.006.
- "†subfamily Dorudontinae Miller 1923 (whale)". Fossilwork.
- "†subfamily Basilosaurinae Miller 1923 (whale)". Fossilworks.
- Gibbes 1847, pp. 8–11; Agassiz 1848, pp. 4–5
- "Basilosaurus". BBC Nature. Retrieved 24 August 2013.
- Harlan 1834.
- "Bry Plantation (Eocene of the United States)". PBDB.
- Switek, Brian (September 21, 2008). "The Legacy of the Basilosaurus". ScienceBlogs. Retrieved 14 September 2013.
- Brian Switek (December 2010). "How Did Whales Evolve?". Smithsonian.
- Harlan 1834, p. 400
- Harlan 1834, p. 401
- Harlan 1834, pp. 402–403
- Owen 1839, pp. 72–73
- Owen 1839, p. 75
- "Wadi Al-Hitan". World Heritage Site. Retrieved 17 August 2017.
- Andrews 1904, pp. 214–215
- Andrews 1906, pp. 255
- Kellogg 1936, p. 81.
- Uhen, Mark D. (2004). "Form, Function, and Anatomy of Dorudon Atrox (Mammalia, Cetacea): An Archaeocete from the Middle to Late Eocene of Egypt". Papers on Paleontology. 34: 11. hdl:2027.42/41255.
- "Fossil Found in Kitchen Counter". National Geographic.
- Basilosauridae in the Paleobiology Database: Taxonomic history. Retrieved August 2013.
- Kellogg 1936, p. 264.
- Zeuglodon vredense (nomen nudum) in the Paleobiology Database. Retrieved August 2013.
- Kellogg 1936, p. 263.
- Zeuglodon puschii (nomen dubium) in the Paleobiology Database. Retrieved August 2013.
- Müller, Johannes Peter (1849). Über die fossilen Reste der Zeuglodonten von Nordamerika mit Rücksicht auf die europäischen Reste aus dieser Familie. Berlin: Reiner. pp. 1–38. OCLC 422134028. Lay summary.
- Uhen, M. D. (2005). "A new genus and species of archaeocete whale from Mississippi". Mississippi Geology. 43 (3): 157–172.
- Uhen, Mark D. (1998). "Middle to Late Eocene Basilosaurines and Dorudontines". The Emergence of Whales. pp. 29–61. doi:10.1007/978-1-4899-0159-0_2. ISBN 978-1-4899-0161-3.
- Basilosaurus drazindai in the Paleobiology Database. Retrieved August 2013.
- Gingerich, Philip D.; Zouhri, Samir (2015). "New fauna of archaeocete whales (Mammalia, Cetacea) from the Bartonian middle Eocene of southern Morocco". Journal of African Earth Sciences. 111: 273–286. Bibcode:2015JAfES.111..273G. doi:10.1016/j.jafrearsci.2015.08.006.
- Gingerich, P. D. (1992). "Marine Mammals (Cetacean and Sirenia) from the Eocene of Gebel Mokattam and Fayum, Egypt: Stratigraphy, Age, and Paleoenvironments". University of Michigan Papers on Paleontology. 30: 1–84. hdl:2027.42/48630. OCLC 26941847.
- Gol'din, Pavel; Zvonok, Evgenij (2013). "Basilotritus uheni, a New Cetacean (Cetacea, Basilosauridae) from the Late Middle Eocene of Eastern Europe". Journal of Paleontology. 87 (2): 254–268. doi:10.1666/12-080R.1.
- Lydekker, R. (2009). "On Zeuglodont and other Cetacean Remains from the Tertiary of the Caucasus". Proceedings of the Zoological Society of London. 1892 (4): 558–581. doi:10.1111/j.1096-3642.1892.tb01782.x.
- "†Microzeuglodon caucasicum Lydekker 1893 (toothed whale)". Fossilworks.
- "†Metasqualodon harwoodi Sanger 1881 (whale)". Fossilworks.org. Retrieved 2019-08-15.
- "On a molar tooth of Zeuglodon from the Tertiary beds on the Murray River near Wellington, S.A." Proceedings of the Linnean Society of New South Wales. 1881.
- Abel, O. (1906). "Ueber den als Beckengurtel von Zeuglodon beschriebenen Schultergurtel eines Vogels aus dem Eocan von Alabama" [On the shoulder girdle of a bird from the Eocene of Alabama described as a pelvic girdle of Zeuglodon]. Centralblatt für Mineralogie, Geologie und Paläontologie. 15: 450–458.
- Lucas, F. A. (1908). "Discussion and Correspondence: is Alabamornis a bird?". Science. 28 (686): 311. doi:10.1126/science.27.686.311. PMID 17770689.
- "State Symbols". State of Mississippi. Archived from the original on 22 August 2013. Retrieved 24 August 2013.
- Fossil whale: State Fossil of Mississippi (PDF), Mississippi Department of Environmental Quality, 1991, retrieved 2019-05-09
- "Sea monster facts". BBC.
- "Walking with Beasts". BBC. Retrieved 9 December 2018.
- "Sea Monsters: A Walking With Dinosaurs Trilogy". BBC.
- Zimmer, Carl (1998). At the Water's Edge: Macroevolution and the Transformation of Life. Free Press. p. 141. ISBN 9780684834900.
Cited literature
- Andrews, C. W. (1904). "Further notes on the mammals of the Eocene of Egypt. Part III". Geological Magazine. 5. 1 (5): 211–215. doi:10.1017/s0016756800119624. Lay summary.
- Andrews, C. W. (1906). A descriptive catalogue of the Tertiary Vertebrata of the Fayûm, Egypt. British Museum (Natural History). OCLC 3675777.
- Brandt, J. F., von (1873). "Über bisher in Russland gefundene Reste von Zeuglodonten". Mélanges Biologiques Tirés du Bulletin de l'Académie Impériale des Sciences de St. Pétersbourg. 9: 111–112. Lay summary (August 2013).
- Harlan, R. (1834). "Notice of fossil bones found in the Tertiary formation of the State of Louisiana". Transactions of the American Philosophical Society. 4: 397–403. doi:10.2307/1004838. JSTOR 1004838. OCLC 63356837.
- Kellogg, R. (1936). A review of the Archaeoceti (PDF, 46.3 Mb). Washington: Carnegie Institution of Washington. OCLC 681376. Publication no. 482.
- Owen, R. (1839). "Observations on the Basilosaurus of Dr. Harlan (Zeuglodon cetoides, Owen)". Transactions of the Geological Society of London. 6: 69–79. doi:10.1144/transgslb.6.1.69. S2CID 84456321. Lay summary.
Further reading
- Gidley, J. W. (1913). "A recently mounted Zeuglodon skeleton in the United States National Museum". Proceedings of the United States National Museum. 44 (1975): 649–654. doi:10.5479/si.00963801.44-1975.649.
- Kellogg, R. (1923). "Description of two squalodonts recently discovered in the Calvert Cliffs, Maryland; and notes on the shark-toothed cetaceans". Proceedings of the United States National Museum. 62 (2462): 1–69. doi:10.5479/si.00963801.62-2462.1. hdl:10088/15278. OCLC 82628874. S2CID 20126705.
- Lucas, Frederic A. (1900). "The pelvic girdle of Zeuglodon, Basilosaurus cetoides (Owen) with notes on other portions of the skeleton". Proceedings of the United States National Museum. 23 (1211): 327–331. doi:10.5479/si.00963801.23-1211.327. hdl:2027/hvd.32044107351454. OCLC 75278090. S2CID 161510206.
- Lydekker, R. (1892). "4. On Zeuglodont and other Cetacean Remains from the Tertiary of the Caucasus". Proceedings of the Zoological Society of London. 60 (4): 558–581. doi:10.1111/j.1096-3642.1892.tb01782.x. OCLC 819196877. Lay summary.
- Sanger, E. B. (1881). "On a molar tooth of Zeuglodon from the Tertiary beds on the Murray River near Wellington, S.A." Proceedings of the Linnean Society of New South Wales. 1 (5): 298–300. doi:10.5962/bhl.part.15884. Archived from the original on 2016-03-06. Lay summary.
External links
| Wikimedia Commons has media related to Basilosaurus. |
Archaeocete genera by family | |||||||
|---|---|---|---|---|---|---|---|
| |||||||
| Pakicetidae | |||||||
| Ambulocetidae | |||||||
| Remingtonocetidae | |||||||
| Protocetidae |
| ||||||
| Basilosauridae |
| ||||||
| |||||||