Coiled coil
A coiled coil is a structural motif in proteins in which 2–7[1] alpha-helices are coiled together like the strands of a rope. (Dimers and trimers are the most common types.) Many coiled coil-type proteins are involved in important biological functions, such as the regulation of gene expression — e.g., transcription factors. Notable examples are the oncoproteins c-Fos and c-jun, as well as the muscle protein tropomyosin.
Discovery
The possibility of coiled coils for α-keratin was initially somewhat controversial. Linus Pauling and Francis Crick independently came to the conclusion that this was possible at about the same time. In the summer of 1952, Pauling visited the laboratory in England where Crick worked. Pauling and Crick met and spoke about various topics; at one point, Crick asked whether Pauling had considered "coiled coils" (Crick came up with the term), to which Pauling said he had. Upon returning to the United States, Pauling resumed research on the topic. He concluded that coiled coils exist, and submitted a lengthy manuscript to the journal Nature in October. Pauling's son Peter Pauling worked at the same lab as Crick, and mentioned the report to him. Crick believed that Pauling had stolen his idea, and submitted a shorter note to Nature a few days after Pauling's manuscript arrived. Eventually, after some controversy and frequent correspondences, Crick's lab declared that the idea had been reached independently by both researchers, and that no intellectual theft had occurred.[2] In his note (which was published first due to its shorter length), Crick proposed the Coiled Coil and as well as mathematical methods for determining their structure.[3] Remarkably, this was soon after the structure of the alpha helix was suggested in 1951 by Linus Pauling and coworkers.[4] These studies were published in the absence of knowledge of a keratin sequence. The first keratin sequences were determined by Hanukoglu and Fuchs in 1982.[5][6]
Based on sequence and secondary structure prediction analyses identified the coiled-coil domains of keratins.[6] These models have been confirmed by structural analyses of coiled-coil domains of keratins.[7]
Molecular structure
Coiled coils usually contain a repeated pattern, hxxhcxc, of hydrophobic (h) and charged (c) amino-acid residues, referred to as a heptad repeat.[8] The positions in the heptad repeat are usually labeled abcdefg, where a and d are the hydrophobic positions, often being occupied by isoleucine, leucine, or valine. Folding a sequence with this repeating pattern into an alpha-helical secondary structure causes the hydrophobic residues to be presented as a 'stripe' that coils gently around the helix in left-handed fashion, forming an amphipathic structure. The most favorable way for two such helices to arrange themselves in the water-filled environment of the cytoplasm is to wrap the hydrophobic strands against each other sandwiched between the hydrophilic amino acids. Thus, it is the burial of hydrophobic surfaces that provides the thermodynamic driving force for the oligomerization. The packing in a coiled-coil interface is exceptionally tight, with almost complete van der Waals contact between the side-chains of the a and d residues. This tight packing was originally predicted by Francis Crick in 1952[3] and is referred to as Knobs into holes packing.
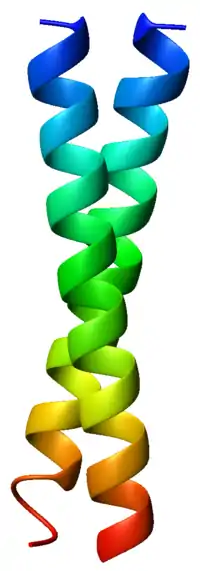
The α-helices may be parallel or anti-parallel, and usually adopt a left-handed super-coil (Figure 1). Although disfavored, a few right-handed coiled coils have also been observed in nature and in designed proteins.[9]
Biological roles
Role in HIV infection
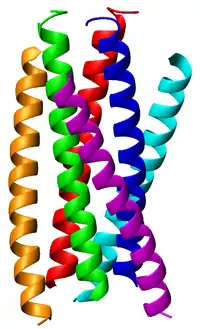
Viral entry into CD4-positive cells commences when three subunits of a glycoprotein 120 (gp120) bind to CD4 receptor and a coreceptor. Glycoprotein gp120 is closely associated to a trimer of gp41 via van der Waals interactions. Upon binding of gp120 to the CD4 receptor and coreceptor, a number of conformational changes in the structure leads to the dissociation of gp120 and to the exposure of gp41 and at the same time to the anchoring of the gp41 N-terminal fusion peptide sequence into the host cell. A spring-loaded mechanism is responsible for bringing the viral and cell membranes in close enough proximity that they will fuse. The origin of the spring-loaded mechanism lies within the exposed gp41, which contains two consecutive heptad repeats (HR1 and HR2) following the fusion peptide at the N terminus of the protein. HR1 forms a parallel, trimeric coiled coil onto which HR2 region coils, forming the trimer-of-hairpins (or six-helix bundle) structure, thereby facilitating membrane fusion through bringing the membranes close to each other. The virus then enters the cell and begins its replication. Recently, inhibitors derived from HR2 such as Fuzeon (DP178, T-20) bind to the HR1 region on gp41 have been developed. However, peptides derived from HR1 have little viral inhibition efficacy due to the propensity for these peptides to aggregate in solution. Chimeras of these HR1-derived peptides with GCN4 leucine zippers have been developed and have shown to be more active than Fuzeon, but these have not entered the clinic yet.
Design
The general problem of deciding on the folded structure of a protein when given the amino acid sequence (the so-called protein folding problem) has not been solved. However, the coiled coil is one of a relatively small number of folding motifs for which the relationships between the sequence and the final folded structure are comparatively well understood.[13][14] Harbury et al. performed a landmark study using an archetypal coiled coil, GCN4, in which rules that govern the way that peptide sequence affects the oligomeric state (that is, the number of alpha-helices in the final assembly) were established.[15][16] The GCN4 coiled coil is a 31-amino-acid (which equates to just over four heptads) parallel, dimeric (i.e., consisting of two alpha-helices) coiled coil and has a repeated isoleucine (or I, in single-letter code) and leucine (L) at the a and d positions, respectively, and forms a dimeric coiled coil. When the amino acids in the a and d positions were changed from I at a and L at d to I at a and I at d, a trimeric (three alpha-helices) coiled coil was formed. Furthermore, switching the positions of L to a and I to d resulted in the formation of a tetrameric (four alpha-helices) coiled coil. These represent a set of rules for the determination of coiled coil oligomeric states and allows scientists to effectively "dial-in" the oligomerization behavior. Another aspect of coiled coil assembly that is relatively well understood, at least in the case of dimeric coiled coils, is that placing a polar residue (in particular asparagine, N) at opposing a positions forces parallel assembly of the coiled coil. This effect is due to a self-complementary hydrogen bonding between these residues, which would go unsatisfied if an N were paired with, for instance, an L on the opposing helix.[17]
It was recently demonstrated by Peacock, Pikramenou and co-workers that coiled coils may be self-assembled using lanthanide(III) ions as a template, thus producing novel imaging agents.[18]
References
- Liu J, Zheng Q, Deng Y, Cheng CS, Kallenbach NR, Lu M (Oct 2006). "A seven-helix coiled coil". Proceedings of the National Academy of Sciences of the United States of America. 103 (42): 15457–62. Bibcode:2006PNAS..10315457L. doi:10.1073/pnas.0604871103. PMC 1622844. PMID 17030805.
- Hager, Thomas. "Narrative 43, Coils Upon Coils". Linus Pauling and the Structure of Proteins. Oregon State University Special Collections and Archives Research Center. Retrieved May 15, 2013.
- Crick FH (Nov 1952). "Is alpha-keratin a coiled coil?". Nature. 170 (4334): 882–3. Bibcode:1952Natur.170..882C. doi:10.1038/170882b0. PMID 13013241. S2CID 4147931.
- Pauling L, Corey RB, Branson HR (Apr 1951). "The structure of proteins; two hydrogen-bonded helical configurations of the polypeptide chain". Proceedings of the National Academy of Sciences of the United States of America. 37 (4): 205–11. Bibcode:1951PNAS...37..205P. doi:10.1073/pnas.37.4.205. PMC 1063337. PMID 14816373.
- Hanukoglu I, Fuchs E (Nov 1982). "The cDNA sequence of a human epidermal keratin: divergence of sequence but conservation of structure among intermediate filament proteins". Cell. 31 (1): 243–52. doi:10.1016/0092-8674(82)90424-X. PMID 6186381. S2CID 35796315.
- Hanukoglu I, Fuchs E (Jul 1983). "The cDNA sequence of a Type II cytoskeletal keratin reveals constant and variable structural domains among keratins". Cell. 33 (3): 915–24. doi:10.1016/0092-8674(83)90034-X. PMID 6191871. S2CID 21490380.
- Hanukoglu I, Ezra L (Jan 2014). "Proteopedia entry: coiled-coil structure of keratins". Biochemistry and Molecular Biology Education. 42 (1): 93–4. doi:10.1002/bmb.20746. PMID 24265184. S2CID 30720797.
- Mason JM, Arndt KM (Feb 2004). "Coiled coil domains: stability, specificity, and biological implications". ChemBioChem. 5 (2): 170–6. doi:10.1002/cbic.200300781. PMID 14760737.
- Harbury PB, Plecs JJ, Tidor B, Alber T, Kim PS (Nov 1998). "High-resolution protein design with backbone freedom". Science. 282 (5393): 1462–7. doi:10.1126/science.282.5393.1462. PMID 9822371.
- Deiss S, Hernandez Alvarez B, Bär K, Ewers CP, Coles M, Albrecht R, Hartmann MD (June 2014). "Your personalized protein structure: Andrei N. Lupas fused to GCN4 adaptors". Journal of Structural Biology. 186 (3): 380–5. doi:10.1016/j.jsb.2014.01.013. PMID 24486584.
- Chou, Hui-Ting; Apelt, Luise; Farrell, Daniel P.; White, Susan Roehl; Woodsmith, Jonathan; Svetlov, Vladimir; Goldstein, Jaclyn S.; Nager, Andrew R.; Li, Zixuan; Muller, Jean; Dollfus, Helene; Nudler, Evgeny; Stelzl, Ulrich; DiMaio, Frank; Nachury, Maxance V.; Walz, Thomas (3 September 2019). "The Molecular Architecture of Native BBSome Obtained by an Integrated Structural Approach". Structure. 27 (9): 1384–1394. doi:10.1016/j.str.2019.06.006. PMC 6726506. PMID 31303482.
- Ludlam, WG; Aoba, T; Cuéllar, J; Bueno-Carrasco, MT; Makaju, A; Moody, JD; Franklin, S; Valpuesta, JM; Willardson, BM (17 September 2019). "Molecular architecture of the Bardet-Biedl syndrome protein 2-7-9 subcomplex". The Journal of Biological Chemistry. 294 (44): 16385–16399. doi:10.1074/jbc.RA119.010150. PMC 6827290. PMID 31530639.
- Bromley EH, Channon K, Moutevelis E, Woolfson DN (Jan 2008). "Peptide and protein building blocks for synthetic biology: from programming biomolecules to self-organized biomolecular systems". ACS Chemical Biology. 3 (1): 38–50. doi:10.1021/cb700249v. PMID 18205291.
- Mahrenholz CC, Abfalter IG, Bodenhofer U, Volkmer R, Hochreiter S (May 2011). "Complex networks govern coiled-coil oligomerization--predicting and profiling by means of a machine learning approach". Molecular & Cellular Proteomics. 10 (5): M110.004994. doi:10.1074/mcp.M110.004994. PMC 3098589. PMID 21311038.
- Harbury PB, Zhang T, Kim PS, Alber T (Nov 1993). "A switch between two-, three-, and four-stranded coiled coils in GCN4 leucine zipper mutants". Science. 262 (5138): 1401–7. Bibcode:1993Sci...262.1401H. doi:10.1126/science.8248779. PMID 8248779. S2CID 45833675.
- Harbury PB, Kim PS, Alber T (Sep 1994). "Crystal structure of an isoleucine-zipper trimer". Nature. 371 (6492): 80–3. Bibcode:1994Natur.371...80H. doi:10.1038/371080a0. PMID 8072533. S2CID 4319206.
- Woolfson, DN (2005). "The design of coiled-coil structures and assemblies". Adv. Protein. Chem. Advances in Protein Chemistry. 70 (4): 79–112. doi:10.1016/S0065-3233(05)70004-8. ISBN 9780120342709. PMID 15837514.
- Berwick MR, Lewis DJ, Jones AW, Parslow RA, Dafforn TR, Cooper HJ, Wilkie J, Pikramenou Z, Britton MM, Peacock AF (Jan 2014). "De novo design of Ln(III) coiled coils for imaging applications". Journal of the American Chemical Society. 136 (4): 1166–9. doi:10.1021/ja408741h. PMC 3950886. PMID 24405157.
Further reading
- Crick, FHC (1953). "The Packing of α-Helices: Simple Coiled-Coils". Acta Crystallogr. 6 (8): 689–697. doi:10.1107/S0365110X53001964.
- Nishikawa K, Scheraga HA (1976). "Geometrical criteria for formation of coiled-coil structures of polypeptide chains". Macromolecules. 9 (3): 395–407. Bibcode:1976MaMol...9..395N. doi:10.1021/ma60051a004. PMID 940353.
- Harbury PB, Zhang T, Kim PS, Alber T (Nov 1993). "A switch between two-, three-, and four-stranded coiled coils in GCN4 leucine zipper mutants". Science. 262 (5138): 1401–7. Bibcode:1993Sci...262.1401H. doi:10.1126/science.8248779. PMID 8248779. S2CID 45833675.
- Gonzalez L, Plecs JJ, Alber T (Jun 1996). "An engineered allosteric switch in leucine-zipper oligomerization". Nature Structural Biology. 3 (6): 510–5. doi:10.1038/nsb0696-510. PMID 8646536. S2CID 30381026.
- Harbury PB, Plecs JJ, Tidor B, Alber T, Kim PS (Nov 1998). "High-resolution protein design with backbone freedom". Science. 282 (5393): 1462–7. doi:10.1126/science.282.5393.1462. PMID 9822371.
- Yu YB (Oct 2002). "Coiled-coils: stability, specificity, and drug delivery potential". Advanced Drug Delivery Reviews. 54 (8): 1113–29. doi:10.1016/S0169-409X(02)00058-3. PMID 12384310.
- Burkhard P, Ivaninskii S, Lustig A (May 2002). "Improving coiled-coil stability by optimizing ionic interactions". Journal of Molecular Biology. 318 (3): 901–10. doi:10.1016/S0022-2836(02)00114-6. PMID 12054832.
- Gillingham AK, Munro S (Aug 2003). "Long coiled-coil proteins and membrane traffic". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1641 (2–3): 71–85. doi:10.1016/S0167-4889(03)00088-0. PMID 12914949.
- Mason JM, Arndt KM (Feb 2004). "Coiled coil domains: stability, specificity, and biological implications". ChemBioChem. 5 (2): 170–6. doi:10.1002/cbic.200300781. PMID 14760737.
External links
Prediction, detection, and visualization
- Spiricoil predict Coiled Coil and Oligormeric state from a protein sequences at Archive.today (archived 2012-12-23)
- NCOILS at Archive.today (archived 2002-01-11)
- Paircoil2 / Paircoil
- bCIPA at the Wayback Machine (archived 2007-02-18)
- STRAP contains an algorithm to predict coiled-coils from AA-sequences.
- PrOCoil predicts the oligomerization of coiled coil proteins and visualizes the contribution of each individual amino acid to the overall oligomeric tendency.
- DrawCoil creates helical wheel diagrams for coiled coils of any oligomerization state and orientation.
Databases
- Spiricoil uses protein domain annotation to predict coiled coil presence and oligormeric state for all completely sequenced organisms
- CC+ is a relational database of coiled coils found in the PDB
- SUPERFAMILY protein domain annotation for all completely sequenced organisms based on the expertly curated SCOP coiled coil class
