Diprotodon
Diprotodon, also known as the Giant wombat, is an extinct genus of diprotodontid marsupial native to Australia during the Pleistocene epoch.
| Diprotodon | |
|---|---|
 | |
| Diprotodon skeleton cast, MNHN, Paris | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Infraclass: | Marsupialia |
| Order: | Diprotodontia |
| Family: | †Diprotodontidae |
| Genus: | †Diprotodon |
| Type species | |
| Diprotodon optatum Owen, 1838 | |
| Synonyms | |
| |
Diprotodon meaning in Greek "two forward teeth".[1] The genus is currently considered monotypic, containing only Diprotodon optatum, the largest known marsupial to have ever existed.[2] It is considered one of the core species of the "Australian megafauna", which ranged throughout the continent during the Pleistocene. Diprotodon existed from about 1.6 million years ago until extinction some 44,000 years ago.[3]
Diprotodon species fossils have been found in sites across mainland Australia,[n 1] including complete skulls, skeletons, and foot impressions.[1] The largest specimens were hippopotamus-sized: about 3 m (9.8 ft) from nose to tail, standing 2 m (6.6 ft) tall at the shoulder and weighing about 2,790 kg (6,150 lb).[5][6][n 2] Diprotodonts may have been depicted on Aboriginal rock art images in Quinkan traditional country (Queensland, Australia).[7][8] Diprotodon became extinct sometime after 44,000 years ago, after the initial settlement of the continent; the role of human and climatic factors in its extinction are uncertain and contested.
The closest surviving relatives of Diprotodon are the wombats and the koala, and so Diprotodon have sometimes been referred to as "giant wombats" in the popular press.[9] Diprotodonts are suggested to have inspired legends of the bunyip, as some Aboriginal tribes identify Diprotodon bones as those of "bunyips".[10]
Discovery
_Diprotodon_australis.png.webp)
The first recorded Diprotodon remains were discovered in a cave near Wellington, New South Wales, in the early 1830s by bushman George Ranken and Major Thomas Mitchell; the latter sent them to England for study by Sir Richard Owen.[11] In the 1840s, Ludwig Leichhardt discovered many Diprotodon bones eroding from the banks of creeks in the Darling Downs of Queensland, and when reporting the find to Owen, commented that the remains were so well preserved, he expected to find living examples in the then-unexplored central regions of Australia.
The majority of fossil finds are of demographic groups indicative of diprotodonts dying in drought conditions. For example, hundreds of individuals were found in Lake Callabonna with well-preserved lower bodies, but crushed and distorted heads.[2] Several family groups are thought to have sunk in mud while crossing the drying lake bed. Other finds consist of age groupings of young or old animals, which are first to die during a drought.
In 2012, a significant group of about 40 was found at Eulo, south-west Queensland.[12]
Taxonomy
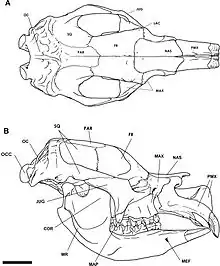
Diprotodon was named by Owen (1838). It was assigned to the Diprotodontidae by McKenna and Bell (1997). The historical classification of Diprotodon consisted of eight species (D. optatum Owen, 1838; D. australis Owen, 1844; D. annextans McCoy, 1861; D. minor Huxley, 1862; D. longiceps McCoy 1865; D. loderi Krefft, 1873a; D. bennettii Krefft, 1873b (nec D. bennettii Owen, 1877); and D. bennettii Owen, 1877 (nec D. bennettii Krefft, 1873b); based on size or slight morphological differences of single specimens collected from isolated geographic regions.[2] Bimodal dental sizes, rather than a continuum of tooth sizes, and identical male and female dental morphology, indicate sexual dimorphism instead of separate species, thus providing strong evidence that the eight species are synonyms for D. optatum.[2] Diprotodon is ultimately thought to have evolved from Euryzygoma dunensis, a smaller diprotodontid known from the Pliocene of eastern Australia, and a smaller form of Diprotodon, labelled D. ?optatum, intermediate in size between the two taxa, is known from the Early Pleistocene (1.77–0.78 Ma) in Nelson Bay near Portland, Victoria.[13]
Description
Diprotodon superficially resembled a rhinoceros without a horn. Its feet turned inwards like a wombat's, giving it a pigeon-toed appearance. It had strong claws on the front feet and its pouch opening faced backwards.[14]
Until recently, how many species of Diprotodon had existed was unknown. Eight species are described, although many researchers believed these actually represented only three at most, while some estimated about 20 in total could exist.
John Walter Gregory collected stories of mystical creatures in Aboriginal myths and legends and considered the possible connections between them and extinct species. He reported a story of a "big, heavy land animal, with a single horn on its forehead" as a possible reference to Diprotodon;[15] the presence of a horn on the rostrum of the species is not scientifically acknowledged.

The skull of Diprotodon has large endocranial sinus cavities, which separate the relatively small cranial vault from the outer part of the skull. These significantly lighten the skull while providing large areas for muscle attachment and reduce load stress.[16]
Paleobiology
Recent research compared the variation between all of the described Diprotodon species with the variation in one of Australia's largest living marsupials, the eastern grey kangaroo, and found the range was comparable, with a near continent-wide distribution.[2] This left only two possible Diprotodon species differing only in size with the smaller being around half the size of the larger. According to Gause's "competitive exclusion principle", no two species with identical ecological requirements can coexist in a stable environment. However, both the small and large diprotodonts coexisted throughout the Pleistocene and the size difference is similar to other sexually dimorphic living marsupials. Further evidence is the battle damage common in competing males found on the larger specimens, but absent from the smaller. Dental morphology also supports sexual dimorphism, with highly sexually dimorphic marsupials, such as the grey kangaroo, having different tooth sizes between males and females, but both sexes having the same dental morphology. An identical dental morphology occurs in the large and small Diprotodon.[2] The taxonomic implication is that Owen's original Diprotodon optatum is the only valid species.
A single sexually dimorphic species allows behavioural interpretations. All sexually dimorphic species of over 5 kg (11 lb) exhibit a polygynous breeding strategy. A modern example of this is the gender segregation of elephants, where females and the young form family groups, while lone males fight for the right to mate with all the females of the group. This behaviour is consistent with fossil finds where adult/juvenile fossil assemblages usually contain only female adult remains.[17][18]
Cyclic variations in the strontium isotope ratios within the tooth enamel of a 300,000-year-old fossil imply that a population of Diprotodon undertook regular, seasonal migrations across the Darling Downs, making it the only known extinct or extant metatherian known to migrate annually.[19] The carbon-13 enamel content was found to have little variation, suggesting a relatively consistent diet through the course of a year containing a mix of both C3 and C4 plants.[19] A finite element method analysis of the skull estimated it had a bite force of around 4500 Newtons at the first molar to over 11,000 N at the fourth molar, values which were described as "exceptionally high", suggesting that Diprotodon was capable of processing tough, fibrous food.[16]
Extinction
Some modern researchers, including Richard Roberts and Tim Flannery, argue that diprotodonts, along with a wide range of other Australian megafauna, became extinct shortly after humans arrived in Australia about 50,000 years ago.[3] Others, including Steve Wroe, note that records in the Australian Pleistocene are rare, and there is not enough data to definitively determine the time of extinction of many of the species, with many of the species having no confirmed record within the last 100,000 years. They suggest that many of the extinctions had been staggered over the course of the late Middle Pleistocene and early Late Pleistocene, prior to human arrival, due to climatic stress.[20] Diprotodon is one of several species with confirmed dates post-dating human arrival on the continent, with the latest high-reliability date being around 44 kyr BP.[21]
Some earlier researchers, including Richard Wright, argued on the contrary that diprotodont remains from several sites, such as Tambar Springs[22] and Trinkey and Lime Springs,[23][24] suggest that Diprotodon survived much longer, into the Holocene. Other more recent researchers, including Lesley Head and Judith Field, favour an extinction date of 28,000 to 30,000 years ago, which would mean that humans coexisted with Diprotodon for some 20,000 years.[25] However, opponents of "late extinction" theories have interpreted such late dates based on indirect dating methods as artifacts resulting from redeposition of skeletal material into more recent strata,[3][26] and recent direct dating results obtained with new technologies have tended to confirm this interpretation.[27][28]
Three theories have been advanced to explain the mass extinction.
Climate change
Australia, as with many other areas of the globe, underwent significant ecological change from predominantly forests to open landscapes during the late Neogene, in correlation with increased cooling. There has been considerable aridification of the Australian interior since the Late Miocene.[29] The recent ice ages produced no significant glaciation in mainland Australia, but long periods of cold and increased aridification. These climatic changes have been suggested as a contributing factor in extinction.[20]
Human hunting
The overkill theory is that human hunters killed and ate the diprotodonts, causing their extinction. The extinctions appear to have coincided with the arrival of humans on the continent. Similar hunting-out happened with the megafauna of New Zealand, Madagascar, and many smaller islands around the world (such as New Caledonia, the Greater Antilles). Critics of this theory regard it as simplistic, arguing that (unlike New Zealand and America), little direct evidence of hunting has been found, and the dates on which the theory rests are too uncertain to be reliable.[20] Linear striated cuts on a Diprotodon tooth from south-eastern Australia initially suggested to have been etched by people are now thought to be bite marks from a spotted‐tailed quoll.[30] A partial juvenile radius of Diprotodon optatum over 47,000 years old was found at the Warratyi rock shelter in South Australia north of Adelaide. Due to the shelter's location on a steep escarpment and the lack of bite marks on the bone, it is thought that humans transported the bone to the site. At present, this represents the only known interaction between humans and Diprotodon.[31]
An examination of swamp sediment cores spanning the last 130,000 years from Lynch's Crater in Queensland suggests that hunting may have been the primary cause of the extinction. Analysis of Sporormiella fungal spores (which are found in the dung of herbivores and are used as a proxy for their abundance) in the cores shows that the Spororniella records in that region virtually disappeared about 41,000 years ago, at a time when climate changes were minimal; the change was accompanied by an increase in charcoal, and was followed by a transition from rainforest to fire-tolerant sclerophyll vegetation.[32] The high-resolution chronology of the changes indicates that fire increased about a century after the disappearance of browsing megafauna, probably due to accumulation of fuel. Grass increased over the next several centuries; sclerophyll vegetation increased following a lag of another century, and a sclerophyll forest developed about a thousand years later.[32] Earlier increases in sclerophyll vegetation during shifts to cooler, drier conditions about 120,000 and 75,000 years ago did not have any obvious impact on megafaunal abundance.[32] A study on Sporormiella in cores going back 135,000 years at the Caledonia Fen wetland in Alpine National Park, found that Sporomiella levels rose after the end of the last interglacial, followed by a sharp drop around 76–60 kya, associated with an increased proportion of wetland plants and further grassland and herbfield; a second sharp drop was observed around 52–45 kya, suggested to represent the megafaunal extinction interval. Charcoal was found throughout the core, suggesting that the arrival of humans and the extinction of megafauna did not change the fire regime at this locality.[33] However, the use of Sporormiella as a megafaunal proxy has been criticised, noting that Sporormiella is found sporadically in the dung of various herbivores, including extant emus and kangaroos, not just megafauna, its presence depends on a variety of factors, often unrelated to megafaunal abundance, and that in Cuddie Springs, a well known megafaunal site, the densities of Sporormiella were consistently low.[34]
Human land management
The third theory says that humans indirectly caused the extinction of diprotodonts by destroying the ecosystem on which they depended. In particular, early Aboriginal people are thought to have been fire-stick farmers using fire regularly and persistently to drive game, open up dense thickets of vegetation, and create fresh green regrowth for both humans and game animals to eat. Evidence for the fire hypothesis is the sudden increase in widespread ash deposits at the time that people arrived in Australia, as well as land-management and hunting practices of modern Aboriginal people as recorded by the earliest European settlers. Evidence against the hypothesis is the fact that humans appear to have eliminated the megafauna of Tasmania without using fire to modify the environment there.[35][36][37]
See also
Notes
- Including Kangaroo Island[4]
- The estimated mean body mass for the 17 adult specimens studied, calculated from the minimum midshaft circumferences of the femur and humerus, is 2,786 kg, although the size of these mammals is believed to be larger because the cartilage is not being accounted for when they fossilize.[6] The 95% confidence interval for this value ranges from 2,272 kg to 3,417 kg.[6]
References
- "Science & Nature: Animals: Diprotodon". Official website. British Broadcasting Corporation (BBC). July 2008. Archived from the original on 2012-02-08. Retrieved 24 February 2011.
- Price, G.J. (June 2008). "Taxonomy and palaeobiology of the largest-ever marsupial, Diprotodon Owen, 1838 (Diprotodontidae, Marsupialia)". Zoological Journal of the Linnean Society. 153 (2): 369–397. doi:10.1111/j.1096-3642.2008.00387.x.
- Roberts, R. G.; Flannery, T. F.; Ayliffe, L. K.; Yoshida, H.; Olley, J. M.; Prideaux, G. J.; Laslett, G. M.; Baynes, A.; Smith, M. A.; Jones, R.; Smith, B. L. (8 June 2001). "New Ages for the Last Australian Megafauna: Continent-Wide Extinction About 46,000 Years Ago" (PDF). Science. 292 (5523): 1888–1892. Bibcode:2001Sci...292.1888R. doi:10.1126/science.1060264. PMID 11397939. S2CID 45643228. Retrieved 26 August 2011.
- Fedorowytsch T.. 2017. Fossil footprints reveal Kangaroo Island's diverse ancient wildlife. ABC Net News. Retrieved on July 24, 2017
- John Pickrell, Ice Age Marsupial Topped Three Tons, Scientists Say, National Geographic News, October 17, 2003.
- Wroe, S.; Crowther, M.; Dortch, J.; Chong, J. (2004-02-07). "The size of the largest marsupial and why it matters". Proceedings of the Royal Society B: Biological Sciences. 271: S34–S36. doi:10.1098/rsbl.2003.0095. PMC 1810005. PMID 15101412.
- "Science notebook - Australian rock painting". The Day. 4 April 1987. Retrieved 7 April 2014.
- Trezise, Percy (1992), Paintings of extinct animals in Quinkan rock art, retrieved 19 November 2019
- Henriques-Gomes, L. (2019-06-27). "'Giant wombat' fossil discovered by council workers in Australia". The Guardian. Retrieved 16 March 2020.
- Shuker, Karl P. N. (1995). "5". In search of prehistoric animals; Do giant extinct creatures still exist? (1st ed.). Blanchford. ISBN 978-0-7137-2469-1.
As far back as 1924, Dr C.W. Anderson of the Australian Museum had suggested that stories of the bunyip could derive from aboriginal legends of the extinct diprotodonts — a view repeated much more recently in Kadimakara (1985) by Australian zoologists Drs. Tim Flannery and Michael Archer, who nominated the palorchestids as plausible candidates.
- Holden, R.; Holden, N. (2001). Bunyips: Australia's Folklore of Fear. National Library Australia. ISBN 978-0642107329.
- Giant marsupials' graveyard unearthed in Queensland By Chrissy Arthur - Australian Broadcasting Corporation - Retrieved 21 July 2012.
- Price, G.J.; Piper, K.J. (December 2009). "Gigantism of the Australian Diprotodon Owen 1838 (Marsupialia, Diprotodontoidea) through the Pleistocene". Journal of Quaternary Science. 24 (8): 1029–1038. doi:10.1002/jqs.1285.
- "Diprotodon | fossil marsupial genus". Encyclopedia Britannica. Retrieved 2020-10-16.
- Gregory, John Walter (1909). The Dead Heart of Australia: A Journey Around Lake Eyre in the Summer of 1901-1902, with Some Account of the Lake Eyre Basin and the Flowing Wells of Central Australia. London: Murray. ISBN 978-0343767174. OCLC 762720327.; pp. 3–7, 74, 224, 230–235.
- Sharp, Alana C.; Rich, Thomas H. (June 2016). "Cranial biomechanics, bite force and function of the endocranial sinuses in Diprotodon optatum, the largest known marsupial". Journal of Anatomy. 228 (6): 984–995. doi:10.1111/joa.12456. PMC 5341585. PMID 26939052.
- Sex secrets of a prehistoric marsupial Archived 2008-06-19 at the Wayback Machine Cosmos Magazine 11 June 2008
- Australasian Science June 2008 Pleistocene Goliath; Gilbert Price
- Price, G. J.; Ferguson, K. J.; Webb, G. E.; Feng, Y.; Higgins, P.; Nguyen, A. D.; Zhao, J.; Joannes-Boyau, R.; Louys, J. (2017). "Seasonal migration of marsupial megafauna in Pleistocene Sahul (Australia–New Guinea)". Proceedings of the Royal Society B: Biological Sciences. 284 (1863): 20170785. doi:10.1098/rspb.2017.0785. PMC 5627191. PMID 28954903.
- Wroe, S.; Field, J. H.; Archer, M.; Grayson, D. K.; Price, G. J.; Louys, J.; Faith, J. T.; Webb, G. E.; Davidson, I.; Mooney, S. D. (2013-05-28). "Climate change frames debate over the extinction of megafauna in Sahul (Pleistocene Australia-New Guinea)". Proceedings of the National Academy of Sciences. 110 (22): 8777–8781. doi:10.1073/pnas.1302698110. ISSN 0027-8424. PMC 3670326. PMID 23650401.
- Johnson, C. N.; Alroy, J.; Beeton, N. J.; Bird, M. I.; Brook, B. W.; Cooper, A.; Gillespie, R.; Herrando-Pérez, S.; Jacobs, Z.; Miller, G. H.; Prideaux, G. J. (2016-02-10). "What caused extinction of the Pleistocene megafauna of Sahul?". Proceedings of the Royal Society B: Biological Sciences. 283 (1824): 20152399. doi:10.1098/rspb.2015.2399. ISSN 0962-8452. PMC 4760161. PMID 26865301.
- Richard Wright (1986). New light on the extinction of the Australian magafauna.
- James Cohen (1995). Aboriginal environmental impacts.
- J.Allen; J.Golson; R.Jones (1977). Sunda and Sahul: Prehistoric Studies in Southeast Asia, Melanesia and Australia. London: Academic Press. pp. 205–246.
- "Future Eaters Ep.1 - Taming the fire -The thesis and responses from critics". Abc.net.au. Retrieved 2014-07-25.
- Flannery, Tim (16 October 2002). The future eaters: an ecological history of the Australasian lands and people. New York: Grove/Atlantic, Inc. ISBN 978-0-8021-3943-6. OCLC 32745413.
- Jones, C. (23 January 2010). "Early humans wiped out Australia's giants". Nature News. doi:10.1038/news.2010.30. Retrieved 7 November 2011.
- Grün, R.; Eggins, S.; Aubert, M.; Spooner, N.; Pike, A. W. G.; Müller, W. (1 March 2010). "ESR and U-series analyses of faunal material from Cuddie Springs, NSW, Australia: implications for the timing of the extinction of the Australian megafauna". Quaternary Science Reviews. 29 (5–6): 596–610. Bibcode:2010QSRv...29..596G. doi:10.1016/j.quascirev.2009.11.004.
- Mao, Xuegang; Retallack, Gregory (January 2019). "Late Miocene drying of central Australia". Palaeogeography, Palaeoclimatology, Palaeoecology. 514: 292–304. doi:10.1016/j.palaeo.2018.10.008.
- Langley, Michelle C. (2020). "Re-analysis of the "engraved" Diprotodon tooth from Spring Creek, Victoria, Australia". Archaeology in Oceania. 55 (1): 33–41. doi:10.1002/arco.5209. ISSN 1834-4453.
- Hamm, G.; Mitchell, P.; Arnold, L.J.; Prideaux, G.J.; Questiaux, D.; .Spooner, N.A.; Levchenko, V.A.; Foley, E.C.; Worthy, T.H.; Stephenson, B.; Coulthard, V. (November 2016). "Cultural innovation and megafauna interaction in the early settlement of arid Australia". Nature. 539 (7628): 280–283. doi:10.1038/nature20125. PMID 27806378. S2CID 4470503.
- Rule, S.; Brook, B. W.; Haberle, S. G.; Turney, C. S. M.; Kershaw, A. P. (2012-03-23). "The Aftermath of Megafaunal Extinction: Ecosystem Transformation in Pleistocene Australia". Science. 335 (6075): 1483–1486. Bibcode:2012Sci...335.1483R. doi:10.1126/science.1214261. PMID 22442481. S2CID 26675232.
- Johnson, Chris N.; Rule, Susan; Haberle, Simon G.; Kershaw, A. Peter; McKenzie, G. Merna; Brook, Barry W. (February 2016). "Geographic variation in the ecological effects of extinction of Australia's Pleistocene megafauna". Ecography. 39 (2): 109–116. doi:10.1111/ecog.01612.
- Dodson, J.; Field, J.H. (May 2018). "What does the occurrence of Sporormiella (Preussia) spores mean in Australian fossil sequences?". Journal of Quaternary Science. 33 (4): 380–392. doi:10.1002/jqs.3020.
- Diamond, Jared (13 August 2008). "Palaeontology: The last giant kangaroo". Nature. 454 (7206): 835–836. Bibcode:2008Natur.454..835D. doi:10.1038/454835a. PMID 18704074. S2CID 36583693.
- Turney, C. S. M.; Flannery, T. F.; Roberts, R. G.; Reid, C.; Fifield, L. K.; Higham, T. F. G.; Jacobs, Z.; Kemp, N.; Colhoun, E. A.; Kalin, R. M.; Ogle, N. (21 August 2008). "Late-surviving megafauna in Tasmania, Australia, implicate human involvement in their extinction". PNAS. NAS. 105 (34): 12150–12153. Bibcode:2008PNAS..10512150T. doi:10.1073/pnas.0801360105. PMC 2527880. PMID 18719103.
- Roberts, R.; Jacobs, Z. (October 2008). "The Lost Giants of Tasmania" (PDF). Australasian Science. 29 (9): 14–17. Archived from the original (PDF) on 27 September 2011. Retrieved 26 August 2011.
External links
| Wikimedia Commons has media related to Diprotodon. |
| Wikispecies has information related to Diprotodon. |
- Australia's lost kingdom on Diprotodon optatum
- BBC science and nature on Diprotodon optatum
- Regional Council of Goyder page on the genera
- Museum Victoria on the Diprotodontids
- Museum Victoria view of a Diprotodon skull
- South Australian Museum information
- Description of Price's research
- 3D model of the skull of Diprotodon
Further reading
- Danielle Clode (2009) Prehistoric giants: the megafauna of Australia. Museum Victoria.
- Barry Cox, Colin Harrison, R.J.G. Savage, and Brian Gardiner. (1999): The Simon & Schuster Encyclopedia of Dinosaurs and Prehistoric Creatures: A Visual Who's Who of Prehistoric Life. Simon & Schuster.
- Jayne Parsons. (2001): Dinosaur Encyclopedia. Dorling Kindersley.
- David Norman. (2001): The Big Book Of Dinosaurs. Welcome Books.
- Gilbert Price. (2005): Article in Memoirs of the Queensland Museum. Queensland Museum.
| |||||||||||||||||||||||
| 
.jpg.webp) | ||||||||||||||||||||||
Vombatomorphia |
| ||||||||||||||||||||||
