Gonadotropin surge-attenuating factor
Gonadotropin surge-attenuating factor (GnSAF) is a nonsteroidal ovarian hormone produced by the granulosa cells of small antral ovarian follicles in females.[1] GnSAF is involved in regulating the secretion of luteinizing hormone (LH) from the anterior pituitary and the ovarian cycle.[1] During the early to mid-follicular phase of the ovarian cycle, GnSAF acts on the anterior pituitary to attenuate LH release, limiting the secretion of LH to only basal levels.[2] At the transition between follicular and luteal phase, GnSAF bioactivity declines sufficiently to permit LH secretion above basal levels, resulting in the mid-cycle LH surge that initiates ovulation.[1] In normally ovulating women, the LH surge only occurs when the oocyte is mature and ready for extrusion.[3] GnSAF bioactivity is responsible for the synchronised, biphasic nature of LH secretion.[4]
Molecular structure and characteristics
GnSAF is a large molecule consisting of subunits that has the same structure as the carboxyl terminal fragment of human serum albumin (HAS).[5][6][7] However, HSA, in its complete form, does not exhibit any GnSAF activity.[8]
The smallest biologically active fraction of GnSAF found in human follicular fluid is a peptide of molecular mass 12.5 kDA.[7] The activity of other subunits has not yet been clarified, but it has been confirmed that more than one protein contributes to the attenuating effect of GnSAF.[9]
Since GnSAF is found in very low concentrations in the human follicular fluid, GnSAF in women has been difficult to isolate, sequence and conclusively characterise.[9]
Synthesis of GnSAF
GnSAF is produced in the granulosa cells of the small sized antral follicles, which have the highest concentration of GnSAF.[10] Concentrations of GnSAF bioactivity is inversely proportional to follicle size.[2] Upon synthesis, GnSAF is released into peripheral circulation.[11]
Follicle-stimulating hormone (FSH) from the anterior pituitary stimulates and prolongs GnSAF biosynthesis in growing small antral follicles in the ovary.[12] FSH induces expression and transcription of exons 12 and 13 of the HSA gene found in granulosa cells.[13] During the early and mid-follicular phase, FSH is secreted to promote growth and proliferation of the granulosa cells, which increases GnSAF concentrations.[14] Once the dominant ovarian follicle has been selected at mid-follicular phase, the non-dominant follicles undergo atresia.[15] Without the presence of small follicles during the late follicular phase, GnSAF concentrations steadily decline to its lowest levels observable in the ovarian cycle.[15] Additionally, the rate of GnSAF biosynthesis by the granulosa cells of the remaining dominant follicle decreases as the follicle approaches maturation.[16] During the transition between luteal phase and follicular phase, GnSAF gradually increases from the late luteal phase and onwards due to the recruitment of follicles and concomitant rise of FSH.[5]
The time-course production of GnSAF depends on the serum FSH concentrations.[17][18] Higher serum concentrations of FSH increases the potency of the attenuating effects of GnSAF on release of LH.[18] However, there is a limit to how much FSH can stimulate GnSAF production: FSH doses above 450 IU do not elicit any more increases in GnSAF bioactivity.[18]
Function
GnSAF antagonises the positive feedback effects of estradiol on GnRH-induced LH release during the follicular phase.[19] GnSAF inhibits the stimulatory role of estradiol in increasing GnRH-induced de novo synthesis of GnRH receptors in the pituitary.[20][21] GnRH receptor mRNA levels are low in the presence of high GnSAF bioactivity, which limits the availability of binding sites for GnRH at the pituitary and decreased pituitary sensitivity to GnRH.[20][21][22] The moderated GnRH pulse amplitude and frequency is sufficient for maintaining low blood concentrations of LH and protects against premature LH surges and LH hypersecretion.[19][23]
GnSAF also inhibits LH synthesis after the transcription stage and limits stored LH in the pituitary.[24] Whilst the pulse amplitude of LH is reduced by GnSAF,[12] constitutive production of LH is unaffected by GnSAF.[25]
Effects on the ovarian cycle in women
Follicular phase
GnSAF prevents a pre-ovulatory surge in LH during this time, allowing sufficient time for the dominant follicle to mature before ovulation.[3][10]
At the start of the follicular phase, high serum FSH concentrations stimulate the development and proliferation of the granulosa cells of the small antral follicles, resulting in a steady increase in GnSAF biosynthesis.[14] The relatively high GnSAF bioactivity dampens the response of the pituitary gland to GnRH by antagonising the sensitising effects of estradiol on the pituitary gland.[22][25] The GnRH pulse, in the presence of GnSAF, is not frequent or potent enough to stimulate LH secretion from the anterior pituitary above basal levels.[26]
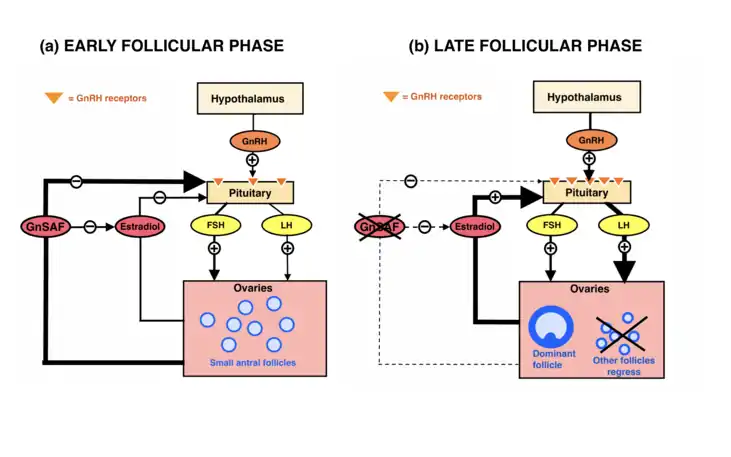
The tonic FSH and LH pulses sufficiently stimulate the theca cells of the follicle to produce androgen substrates for granulosa cell aromatase and induces cytochrome P450 enzymes that can produce progesterone later in the luteal phase.[3] Androgen substrates from the theca cells are used by the enlarging granulosa cells to produce more estradiol.[3]
At mid-follicular phase, GnSAF bioactivity gradually declines as the dominant follicle is established and the small subordinate follicles undergo atresia.[15] The development of the dominant follicle and regression of small non-dominant follicles is supported by increasing estradiol secretion.[15]
Towards the end of the follicular phase, GnSAF bioactivity is at its lowest due to the absence of small antral follicles.[1] Estradiol secretion from the dominant follicle exponentially increases and exceeds a threshold which switches estradiol feedback on GnRH pulse frequency from negative to positive.[12][15] Pituitary sensitivity to GnRH is restored.[2][4]
Mid-cycle
The absence of GnSAF lowers the threshold for GnRH pulse frequency and amplitude required to stimulate the anterior pituitary to secrete LH.[19] Concurrently, GnRH pulse frequency and amplitude increases, which allows LH secretion significantly higher than basal levels.[23] The LH secretion most noticeable as a LH surge lasting 48 to 72 hours in the middle of the ovarian cycle.[3] Meiosis in the dominant follicle resumes and follicle ruptures shortly after the LH surge.[27] Ovulation can only occur if GnSAF is absent and the mid-cycle LH surge occurs.[3][27]
Luteal phase
Immediately after the LH surge and subsequent ovulation, estradiol concentrations drop and the corpus luteum develops.[23]
Towards the end of luteal phase, GnSAF production in the small antral follicles increases steadily.[18] FSH does not stimulate GnSAF production in the corpus luteum, so GnSAF bioactivity is low after ovulation, until the intercycle rise in FSH occurs.[16]
Relationship between GnRH and GnSAF
GnRH and GnSAF are functionally antagonistic over the control of LH secretion in the hypothalamic-pituitary axis.[15] In the presence of GnSAF, endogenous pulses of GnRH from the hypothalamus still persist, in approximately one hour intervals.[23] Due to this large time interval between consecutive GnRH pulses, GnSAF effectively limits the effects of GnRH on the anterior pituitary.[23] GnSAF acts on the gonadotropic cells of the pituitary to neutralise the second messenger pathway responsible for transducing GnRH signalling in the gonadotropes.[28] The effectiveness of downstream actions of GnRH, such as calcium mobilisation and the protein kinase C system, are reduced by GnSAF.[28] These antagonistic effects of GnSAF on GnRH keeps the anterior pituitary in a low responsiveness state, which prevents acute elevations of serum LH concentrations until GnSAF bioactivity declines.[28]
When estradiol concentrations are high in the late follicular phase, GnRH pulse frequency and amplitude increases and overrides the attenuating effects of GnSAF.[23] Frequent and consecutive exogenous administration of GnRH at submaximal doses is sufficient in overcoming the neutralizing effects of GnSAF.[29] This is because estradiol lowers the GnRH pulse frequency and amplitude required to stimulate the biosynthesis and secretion of LH.[30]
Associated disease states and abnormalities
Abnormal GnSAF bioactivity has been associated with premature surges in LH and LH hypersecretion. Optimal and timely changes in serum LH concentrations are crucial to ensuring the viability of oocytes and implantation after fertilization.[15][23] For successful implantation of a zygote, the mid-cycle LH surge after the decline of GnSAF and ovulation must correspond with uterine receptivity.[23] Hypersecretion of LH contributes to cycle disturbance, infertility and increased chances of miscarriage.[31]
GnSAF has been implicated in polycystic ovary syndrome (PCOS), one of the most common ovarian disorders responsible for causing anovulatory infertility.[32] Approximately 40% of women with PCOS display higher GnRH pulse frequency and tonic hypersecretion of LH due to hypersecretion of androgens from the polycystic ovary.[3][32][33] Androgens are readily metabolized to estradiol in the ovaries.[33] The supraphysiological concentrations of estradiol maintains high pituitary responsiveness to GnRH, permitting the hypersecretion of LH.[34]
Superovulation is common in women who take medications such as clomiphene citrate, an anti-estrogenic oral medication used to treat infertility.[22] Superovulation is induced in women to increase chances of fertilization and conception in assisted reproductive techniques.[22] In naturally superovulating women, the mid-cycle LH surge is significantly lower compared to that of normal ovulating women due to the presence of GnSAF in the late follicular phase fluid.[17][35] The high basal GnSAF bioactivity is due to a greater number of small follicles, the site of GnSAF biosynthesis.[36]
Possible uses in medicine
An alternative to using GnRH analogues in IVF treatments could be short-term administration of GnSAF.[37] During IVF, the ovaries are stimulated by raising estrogen concentrations to supraphysiological levels, which prevents the mid-cycle LH surge.[22][35] Premature LH surges are unfavorable during IVF as it is associated with low oocyte viability and low success rates during IVF treatment.[22] GnSAF could be used to influence ovarian hyperstimulation syndrome.[38] Using GnSAF would potentially eliminate the need to use human chorionic gonadotropin.[38]
Administration of GnSAF could also be used to prevent ovulation and replace exogenously administered steroids that are often perceived as being risky,[39] or to delay the naturally premature LH surge observed in some hyperstimulated or infertile women.[15] GnSAF could form part of a contraceptive drug[37] or in treatments for infertility that target LH hypersecretion or abnormal ovarian cycles.[15]
References
- Messinis, Ioannis E.; Messini, Christina I.; Anifandis, George; Garas, Antonios; Daponte, Alexandros (2018), "Gonadotropin Surge-Attenuating Factor: A Nonsteroidal Ovarian Hormone Controlling GnRH-Induced LH Secretion in the Normal Menstrual Cycle", Vitamins and Hormones, Elsevier, 107: 263–286, doi:10.1016/bs.vh.2018.01.002, ISBN 9780128143599, PMID 29544633
- Fowler, P. A. (2001-07-01). "Relationship between follicle size and gonadotrophin surge attenuating factor (GnSAF) bioactivity during spontaneous cycles in women". Human Reproduction. 16 (7): 1353–1358. doi:10.1093/humrep/16.7.1353. ISSN 1460-2350. PMID 11425812.
- Wallach, Edward E.; Shoham, Zeev; Schachter, Morey; Loumaye, Ernest; Weissman, Ariel; MacNamee, Michel; Insler, Vaclav (1995). "The luteinizing hormone surge—the final stage in ovulation induction: modern aspects of ovulation triggering". Fertility and Sterility. 64 (2): 237–251. doi:10.1016/s0015-0282(16)57717-6. ISSN 0015-0282. PMID 7615097.
- Messinis, I.E.; Lolis, D.; Zikopoulos, K.; Tsahalina, E.; Seferiadis, K.; Templeton, A.A. (1994). "Modulation of the action of gonadotrophin surge-attenuating factor by gonadotrophin-releasing hormone". Human Reproduction. 9 (8): 1437–1441. doi:10.1093/oxfordjournals.humrep.a138725. ISSN 1460-2350. PMID 7989501.
- MESSINIS, IOANNIS E. (2003). "Modulatory Effect of the Ovary on LH Secretion". Annals of the New York Academy of Sciences. 997 (1): 35–41. Bibcode:2003NYASA.997...35M. doi:10.1196/annals.1290.004. ISSN 0077-8923.
- Danforth, D. R.; Cheng, C. Y. (1993). "The identification of gonadotrophin surge inhibiting factor and its role in the regulation of pituitary gonadotrophin secretion". Human Reproduction. 8 (suppl 2): 117–122. doi:10.1093/humrep/8.suppl_2.117. ISSN 0268-1161.
- Pappa, Aglaia; Seferiadis, Konstantin; Fotsis, Theodore; Shevchenko, Andrej; Marselos, Marios; Tsolas, Orestes; E.Messinis, Ioannis (1999). "Purification of a candidate gonadotrophin surge attenuating factor from human follicular fluid". Human Reproduction. 14 (6): 1449–1456. doi:10.1093/humrep/14.6.1449. ISSN 1460-2350.
- Tavoulari, S. (2004). "The recombinant subdomain IIIB of human serum albumin displays activity of gonadotrophin surge-attenuating factor". Human Reproduction. 19 (4): 849–858. doi:10.1093/humrep/deh187. ISSN 1460-2350. PMID 15016777.
- Fowler, P. A. (2002). "A 60-66 kDa protein with gonadotrophin surge attenuating factor bioactivity is produced by human ovarian granulosa cells". Molecular Human Reproduction. 8 (9): 823–832. doi:10.1093/molehr/8.9.823. ISSN 1460-2407. PMID 12200460.
- Fowler, P. A.; Messinis, I. E.; Templeton, A. A. (1990). "Inhibition of LHRH-induced LH and FSH release by gonadotrophin surge-attenuating factor (GnSAF) from human follicular fluid". Reproduction. 90 (2): 587–594. doi:10.1530/jrf.0.0900587. ISSN 1470-1626. PMID 2123501.
- Fowler, P A; Cunningham, P; Fraser, M; MacGregor, F; Byrne, B; Pappas, A; Messinis, I E; Templeton, A (1994). "Circulating gonadotrophin surge-attenuating factor from superovulated women suppresses in vitro gonadotrophin-releasing hormone self-priming". Journal of Endocrinology. 143 (1): 45–54. doi:10.1677/joe.0.1430045. ISSN 0022-0795. PMID 7964321.
- Fowler, P. (2003-12-01). "Ovarian gonadotrophin surge-attenuating factor (GnSAF): where are we after 20 years of research?". Reproduction. 126 (6): 689–699. doi:10.1530/reprod/126.6.689. ISSN 1470-1626.
- Karligiotou, E.; Kollia, P.; Kallitsaris, A.; Messinis, I.E. (2005-10-27). "Expression of human serum albumin (HSA) mRNA in human granulosa cells: potential correlation of the 95 amino acid long carboxyl terminal of HSA to gonadotrophin surge-attenuating factor". Human Reproduction. 21 (3): 645–650. doi:10.1093/humrep/dei374. ISSN 1460-2350. PMID 16253963.
- Messinis, I. E.; Hirsch, P.; Templeton, A. A. (1991). "Follicle stimulating hormone stimulates the production of gonadotrophin surge attenuating factor (GnSAF) in vivo". Clinical Endocrinology. 35 (5): 403–407. doi:10.1111/j.1365-2265.1991.tb03556.x. ISSN 0300-0664. PMID 1814653.
- Fowler, P. A. (1996-04-01). "The nature and function of putative gonadotropin surge- attenuating/inhibiting factor (GnSAF/IF)". Endocrine Reviews. 17 (2): 103–120. doi:10.1210/er.17.2.103. ISSN 0163-769X.
- Messinis, Ioannis E.; Messini, Christina I.; Dafopoulos, Konstantinos (2014). "Novel aspects of the endocrinology of the menstrual cycle". Reproductive BioMedicine Online. 28 (6): 714–722. doi:10.1016/j.rbmo.2014.02.003. ISSN 1472-6483. PMID 24745832.
- MESSINIS, IOANNIS E.; TEMPLETON, ALLAN (1990). "IN-VIVO BIOACTIVITY OF GONADOTROPHIN SURGE ATTENUATING FACTOR (GnSAF)". Clinical Endocrinology. 33 (2): 213–218. doi:10.1111/j.1365-2265.1990.tb00485.x. ISSN 0300-0664. PMID 2121393.
- Messinis, I. E.; Lolis, D.; Papadopoulos, L.; Tsahallna, Th.; Papanikolaou, N.; Seferiadis, K.; Templeton, A. A. (1993). "Effect of varying concentrations of follicle stimulating hormone on the production of gonadotrophin surge attenuating factor (GnSAF) in women". Clinical Endocrinology. 39 (1): 45–50. doi:10.1111/j.1365-2265.1993.tb01749.x. ISSN 0300-0664.
- Fowler, P.A.; Messinis, I.E.; Cunningham, P.; Fraser, M.; Templeton, A.A. (1993). "Effects of gonadotrophin surge-attenuating factor on the two pools of gonadotrophin-releasing hormone-induced luteinizing hormone secretion in vitro". Human Reproduction. 8 (6): 822–828. doi:10.1093/oxfordjournals.humrep.a138148. ISSN 1460-2350. PMID 8345069.
- Menon, M.; Peegel, H.; Katta, V. (1985). "Estradiol potentiation of gonadotropin-releasing hormone responsiveness in the anterior pituitary is mediated by an increase in gonadotropin-releasing hormone receptors". American Journal of Obstetrics and Gynecology. 151 (4): 534–540. doi:10.1016/0002-9378(85)90284-4. ISSN 0002-9378. PMID 2983554.
- AIYER, M. S.; CHIAPPA, SHARON A.; FINK, G. (1974). "A Priming Effect of Luteinizing Hormone Releasing Factor on the Anterior Pituitary Gland in the Female Rat". Journal of Endocrinology. 62 (3): 573–588. doi:10.1677/joe.0.0620573. ISSN 0022-0795. PMID 4606132.
- Messinis, Ioannis E. (2006). "Ovarian feedback, mechanism of action and possible clinical implications". Human Reproduction Update. 12 (5): 557–571. doi:10.1093/humupd/dml020. ISSN 1460-2369. PMID 16672246.
- de Koning, J. (2001-02-01). "Is GnRH self-priming an obligatory feature of the reproductive cycle?". Human Reproduction. 16 (2): 209–214. doi:10.1093/humrep/16.2.209. ISSN 1460-2350. PMID 11157808.
- Gordon, Ana; Garrido-Gracia, Jose C.; Aguilar, Rafaela; Sánchez-Criado, José E. (2009). "Ovarian stimulation with FSH in the rat reduces proestrous GnRH-dependent LH secretion through a dual mechanism: Inhibition of LH synthesis and release". Neuroscience Letters. 460 (3): 219–222. doi:10.1016/j.neulet.2009.05.055. ISSN 0304-3940. PMID 19477224.
- Bolen, Adam H.; Jacobs, Howard S. (1991). "Gonadotrophin Surge Attenuating Factor: a missing link in the control of LH secretion?". Clinical Endocrinology. 35 (5): 399–402. doi:10.1111/j.1365-2265.1991.tb03555.x. ISSN 0300-0664. PMID 1814652.
- Lüking Jayes, Friederike C.; Britt, Jack H.; Esbenshade, Kenneth L. (1997). "Role of Gonadotropin-Releasing Hormone Pulse Frequency in Differential Regulation of Gonadotropins in the Gilt". Biology of Reproduction. 56 (4): 1012–1019. doi:10.1095/biolreprod56.4.1012. ISSN 0006-3363. PMID 9096885.
- Liu, Lei; Kong, Nana; Xia, Guoliang; Zhang, Meijia (2013). "Molecular control of oocyte meiotic arrest and resumption". Reproduction, Fertility and Development. 25 (3): 463–71. doi:10.1071/rd12310. ISSN 1031-3613. PMID 23217677.
- Tijssen, A. M. I.; Helder, M. N.; Chu, Z-W.; de Koning, J. (1997). "Intracellular antagonistic interaction between GnRH and gonadotrophin surge-inhibiting/attenuating factor bioactivity downstream of second messengers involved in the self-priming process". Reproduction. 111 (2): 235–242. doi:10.1530/jrf.0.1110235. ISSN 1470-1626. PMID 9462291.
- Messinis, I. E.; Lolis, D.; Zikopoulos, K.; Milingos, S.; Kollios, G.; Seferiadis, K.; Templeton, A. A. (1996). "Effect of follicle stimulating hormone or human chorionic gonadotrophin treatment on the production of gonadotrophin surge attenuating factor (GnSAF) during the luteal phase of the human menstrual cycle". Clinical Endocrinology. 44 (2): 169–175. doi:10.1046/j.1365-2265.1996.589411.x. ISSN 0300-0664. PMID 8849571.
- RAMEY, J. W.; HIGHSMITH, R. F.; WILFINGER, W. W.; BALDWIN, D. M. (1987). "The Effects of Gonadotropin-Releasing Hormone and Estradiol on Luteinizing Hormone Biosynthesis in Cultured Rat Anterior Pituitary Cells*". Endocrinology. 120 (4): 1503–1513. doi:10.1210/endo-120-4-1503. ISSN 0013-7227.
- Homburg, R; Armar, NA; Eshel, A; Adams, J; Jacobs, HS (1989). "Influence of serum luteinising hormone concentrations on ovulation, conception, and early pregnancy loss in polycystic ovary syndrome". International Journal of Gynecology & Obstetrics. 28 (4): 1024–1026. doi:10.1016/0020-7292(89)90625-5. ISSN 0020-7292. PMC 1834779. PMID 3142595.
- Balen, A. H. (1993-11-01). "Hypersecretion of luteinizing hormone and the polycystic ovary syndrome". Human Reproduction. 8 (suppl 2): 123–128. doi:10.1093/humrep/8.suppl_2.123. ISSN 0268-1161. PMID 8276945.
- Balen, Adam H. (2001), "In vitro fertilization and the patient with polycystic ovaries or polycystic ovary syndrome", Polycystic Ovary Syndrome, Cambridge University Press, pp. 177–203, doi:10.1017/cbo9780511545191.012, ISBN 9780511545191
- WANG, C. F.; LASLEY, B. L.; LEIN, A.; YEN, S. S. C. (1976). "The Functional Changes of the Pituitary Gonadotrophs During the Menstrual Cycle". The Journal of Clinical Endocrinology & Metabolism. 42 (4): 718–728. doi:10.1210/jcem-42-4-718. ISSN 0021-972X. PMID 770496.
- MESSINIS, IOANNIS E.; TEMPLETON, ALLAN; BAIRD, DAVID T. (1985). "Endogenous Luteinizing Hormone Surge during Superovulation Induction with Sequential Use of Clomiphene Citrate and Pulsatile Human Menopausal Gonadotropin". The Journal of Clinical Endocrinology & Metabolism. 61 (6): 1076–1080. doi:10.1210/jcem-61-6-1076. ISSN 0021-972X.
- MESSINIS, IOANNIS E.; TEMPLETON, ALLAN (1990). "Superovulation Induction in Women Suppresses Luteinizing Hormone Secretion at the Pituitary Level". Clinical Endocrinology. 32 (1): 107–114. doi:10.1111/j.1365-2265.1990.tb03756.x. ISSN 0300-0664.
- Vega, Mario G.; Zarek, Shvetha M.; Bhagwat, Medha; Segars, James H. (2015). "Gonadotropin surge-inhibiting/attenuating factors: A review of current evidence, potential applications, and future directions for research". Molecular Reproduction and Development. 82 (1): 2–16. doi:10.1002/mrd.22439. ISSN 1040-452X. PMC 4710370. PMID 25581424.
- Delvigne, A. (2003-01-01). "Review of clinical course and treatment of ovarian hyperstimulation syndrome (OHSS)". Human Reproduction Update. 9 (1): 77–96. doi:10.1093/humupd/dmg005. ISSN 1355-4786. PMID 12638783.
- HODGEN, GARY D.; KENIGSBURG, DANIEL; COLLINS, ROBERT L.; SCHENKEN, ROBERT S. (1985). "Selection of the Dominant Ovarian Follicle and Hormonal Enhancement of the Natural Cycle". Annals of the New York Academy of Sciences. 442 (1 In Vitro Fert): 23–37. Bibcode:1985NYASA.442...23H. doi:10.1111/j.1749-6632.1985.tb37502.x. ISSN 0077-8923. PMID 3925837.