Haplogroup H (mtDNA)
Haplogroup H is a human mitochondrial DNA (mtDNA) haplogroup. The clade is believed to have originated in Southwest Asia, near present day Syria,[1] around 20,000 to 25,000 years ago. Mitochondrial haplogroup H is today predominantly found in Europe, and is believed to have evolved before the Last Glacial Maximum (LGM). It first expanded in the northern Near East and Southern Caucasus soon, and later migrations from Iberia suggest that the clade reached Europe before the Last Glacial Maximum. The haplogroup has also spread to parts of Africa, Siberia and inner Asia. Today, around 40% of all maternal lineages in Europe belong to haplogroup H.
| Haplogroup H | |
|---|---|
| Possible time of origin | 20,000–25,000 YBP |
| Possible place of origin | Southwest Asia[1] |
| Ancestor | HV[1] |
| Descendants | H* lineages; subclades H1, H2, H3, H4, H5'36, H6, H7, H8, H9, H10, H11, H12, H13, H14, H15, H16, H18, H19, H20, H22, H23, H24, H25, H26, H28, H29, H31, H32, H33, H34, H35, H37, H38, H39, H53, 16129(H17+H27), 16129(H21+H30) (numbers upto H135)[2] |
| Defining mutations | G2706A, T7028C[3] |
Origin
Haplogroup H is a descendant of haplogroup HV. The Cambridge Reference Sequence (CRS), which until recently was the human mitochondrial sequence to which all others were compared, belongs to haplogroup H2a2a1 (human mitochondrial sequences should now be compared with the ancestral Reconstructed Sapiens Reference Sequence (RSRS)).[4] Several independent studies conclude that haplogroup H probably evolved in West Asia c. 25,000 years ago.
In July 2008 ancient mtDNA from an individual called Paglicci 23, whose remains were dated to 28,000 years ago and excavated from Paglicci Cave (Apulia, Italy), were found to be identical to the Cambridge Reference Sequence in HVR1.[5] This once was believed to indicate haplogroup H, but researchers now recognize that CRS HVR1 also appears in U or HV, because there are no HVR1 mutations that separate CRS from the haplogroup R founder. Haplogroup HV derives from the haplogroup R0 which in turn derives from haplogroup R is a descendant of macro-haplogroup N like its sibling M, is a descendant of haplogroup L3.
Haplogroup H has also been found among Iberomaurusian specimens dating from the Epipaleolithic at the Taforalt and Afalou prehistoric sites.[6] Among the Taforalt individuals, around 29% of the observed haplotypes belonged to various H subclades, including H1 (2/24; 8%), H103 (1/24; 4%), H14b1 (1/24; 4%), H2a2a1 (1/24; 4%), and H2a1e1a (1/24; 4%). A further 41% of the analysed haplotypes could be assigned to either haplogroup H or haplogroup U. Among the Afalou individuals, the H subclades were represented by H103 (1/9; 11%). A further 44% of the analysed haplotypes could be assigned to either haplogroup H or haplogroup U (3/9; 33%) or to haplogroup H14b1 or haplogroup JT (1/9; 11%).[7]
MtDNA H had frequency of 19% among Neolithic Early European Farmers and virtually absent among Mesolithic European hunter gatherers.[8]
MtDNA H was also present among Trypillians.[9]
The clade has been observed among ancient Egyptian mummies excavated at the Abusir el-Meleq archaeological site in Middle Egypt, which date from the Pre-Ptolemaic/late New Kingdom and Ptolemaic periods.[10]
Additionally, haplogroup H has been found among specimens at the mainland cemetery in Kulubnarti, Sudan, which date from the Early Christian period (AD 550–800).[11]
Distribution
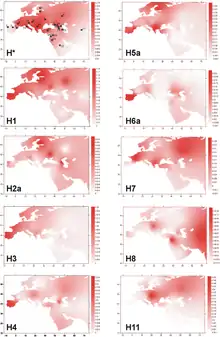
Haplogroup H is the most common mtDNA clade in Europe.[12] It is found in approximately 41% of native Europeans.[13][14] The lineage is also common in North Africa and the Middle East.[15]
The majority of the European populations have an overall haplogroup H frequency of 40–50%, with frequencies decreasing in the southeast. The clade reaches 20% in the Near East and Caucasus, 17% in Iran, and <10% in the Arabian Peninsula, Northern India and Central Asia.[1][16]
Undifferentiated haplogroup H has been found among Palestinians (14%),[17] Syrians (13.6%),[17] Druze (10.6%),[17] Iraqis (9.5%),[17] Somalis (6.7%),[17] Egyptians (5.7% in El-Hayez;[18] 14.7% in Gurna[19]), Saudis (5.3–10%),[17] Soqotri (3.1%),[20] Nubians (1.3%),[17] and Yemenis (0–13.9%).[17]
Subclades
Among all these clades, the subhaplogroups H1 and H3 have been subject to a more detailed study and would be associated to the Magdalenian expansion from SW Europe c. 13,000 years ago:[21]
H1
_of_haplogroup_H1_in_western_Eurasia_and_North_Africa..png.webp)
H1 encompasses an important fraction of Western European mtDNA lineages, reaching its local peak among contemporary Basques (27.8%). The clade also occurs at high frequencies elsewhere in the Iberian Peninsula, as well as in the Maghreb (Tamazgha). The haplogroup frequency is above 10% in many other parts of Europe (France, Sardinia, parts of the British Isles, Alps, large portions of Eastern Europe), and surpasses 5% in nearly all of the continent.[1] Its H1b subclade is most common in eastern Europe and NW Siberia.[22]
As of 2010, the highest frequency of the H1 subclade has been found among the Tuareg inhabiting the Fezzan region in Libya (61%).[23] The basal H1* haplogroup is found among the Tuareg inhabiting the Gossi area in Mali (4.76%).[24]
The rare H1cb subclade is concentrated among Fulani groups inhabiting the Sahel.[25]
Haplogroup H has been found in various fossils that were analysed for ancient DNA, including specimens associated with the Linearbandkeramik culture (H1e, Halberstadt-Sonntagsfeld, 1/22 or ~5%; H1 or H1au1b, Karsdorf, 1/2 or 50%), Germany Middle Neolithic (H1e1a, Esperstedt, 1/1 or 100%), Iberia Early Neolithic (H1, El Prado de Pancorbo, 1/2 or 50%), Iberia Middle Neolithic (H1, La Mina, 1/4 or 25%), and Iberia Chalcolithic (H1t, El Mirador Cave, 1/12 or ~8%).[26] Haplogroup H has been observed in ancient Guanche fossils excavated in Gran Canaria and Tenerife on the Canary Islands, which have been radiocarbon-dated to between the 7th and 11th centuries CE. At the Tenerife site, these clade-bearing individuals were found to belong to the H1cf subclade (1/7; ~14%); at the Gran Canaria site, the specimens carried the H2a subhaplogroup (1/4; 25%).[27] Additionally, ancient Guanche (Bimbaches) individuals excavated in Punta Azul, El Hierro, Canary Islands were all found to belong to the H1 maternal subclade. These locally born individuals were dated to the 10th century and carried the H1-16260 haplotype, which is exclusive to the Canary Islands and Algeria.[28]
- Frequencies of haplogroup H1 in the world (Ottoni et al. 2010)
| Region or Population | H1% | No. of subjects |
|---|---|---|
| Africa | ||
| Libyan Tuareg | 61 | 129 |
| Tuareg (West Sahel) | 23.3 | 90 |
| Berbers (Morocco) | 20.2 | 217 |
| Morocco | 12.2 | 180 |
| Berbers (Tunisia) | 13.4 | 276 |
| Tunisia | 10.6 | 269 |
| Mozabite | 9.8 | 80 |
| Siwas (Egypt) | 1.1 | 184 |
| Western Sahara | 14.8 | 128 |
| Mauritania | 6.9 | 102 |
| Senegal | 0 | 100 |
| Fulani (Chad–Cameroon) | 0 | 186 |
| Cameroon | 0 | 142 |
| Chad | 0 | 77 |
| Buduma (Niger) | 0 | 30 |
| Nigeria | 0 | 69 |
| Ethiopia | 0 | 82 |
| Amhara (Ethiopia) | 0 | 90 |
| Oromo (Ethiopia) | 0 | 117 |
| Sierra Leone | 0 | 155 |
| Guineans (Guiné Bissau) | 0 | 372 |
| Mali | 0 | 83 |
| Kikuyu (Kenya) | 0 | 24 |
| Benin | 0 | 192 |
| Asia | ||
| Central Asia | 0.7 | 445 |
| Pakistan | 0 | 100 |
| Yakuts | 1.7 | 58 |
| Caucasus | ||
| Caucasus (north) | 8.8 | 68 |
| Caucasus (south) | 2.3 | 132 |
| Northwestern Caucasus | 4.7 | 234 |
| Armenians | 2.3 | 175 |
| Daghestan | 2.5 | 269 |
| Georgians | 1 | 193 |
| Karachay-Balkars | 4.4 | 203 |
| Ossetians | 2.4 | 296 |
| Europe | ||
| Andalusia | 24.3 | 103 |
| Basques (Spain) | 27.8 | 108 |
| Catalonia | 13.9 | 101 |
| Galicia | 17.7 | 266 |
| Pasiegos (Cantabria) | 23.5 | 51 |
| Portugal | 25.5 | 499 |
| Spain (miscellaneous) | 18.9 | 132 |
| Italy (north) | 11.5 | 322 |
| Italy (center) | 6.3 | 208 |
| Italy (south) | 8.7 | 206 |
| Sardinia | 17.9 | 106 |
| Sicily | 10 | 90 |
| Finland | 18 | 78 |
| Volga-Ural Finnic speakers | 13.6 | 125 |
| Basques (France) | 17.5 | 40 |
| Béarnaise | 14.8 | 27 |
| France | 12.3 | 106 |
| Estonia | 16.7 | 114 |
| Saami | 0 | 57 |
| Lithuania | 1.7 | 180 |
| Hungary | 11.3 | 303 |
| Czech Republic | 10.8 | 102 |
| Ukraine | 9.9 | 191 |
| Poland | 9.3 | 86 |
| Russia | 13.5 | 312 |
| Austria | 10.6 | 2487 |
| Germany | 6 | 100 |
| Romania | 9.4 | 360 |
| Netherlands | 8.8 | 34 |
| Greece (Aegean islands) | 1.6 | 247 |
| Greece (mainland) | 6.3 | 79 |
| Macedonia | 7.1 | 252 |
| Albania | 2.9 | 105 |
| Turks | 3.3 | 360 |
| Balkans | 5.4 | 111 |
| Croatia | 8.3 | 84 |
| Slovaks | 7.6 | 119 |
| Slovak (East) | 16.8 | 137 |
| Slovak (West) | 14.2 | 70 |
| Middle East | ||
| Arabian Peninsula | 0 | 94 |
| Arabian Peninsula (incl. Yemen, Oman) | 0.8 | 493 |
| Druze | 3.4 | 58 |
| Dubai (United Arab Emirates) | 0.4 | 249 |
| Iraq | 1.9 | 206 |
| Jordanians | 1.7 | 173 |
| Lebanese | 4.2 | 167 |
| Syrians | 0 | 159 |
H3
H3 is found throughout the whole of Europe and in the Maghreb but does not exist in the Far East ,[1] and is believed to have originated among Mesolithic hunter-gatherers in south-western Europe between 9 000 and 11 000 years ago. H3 represents the second largest fraction of the H genome after H1 and has a somewhat similar distribution, with peaks in Portugal, Spain, Scandinavia and Finland. It is common in Portugal (12%), Sardinia (11%), Galicia (10%), the Basque country (10%), Ireland (6%), Norway (6%), Hungary (6%) and southwestern France (5%).[1][29][30] Studies have suggested haplogroup H3 is highly protective against AIDS progression.[31]
Example of H3 sub-groups are:[30]
- H3a and H3g, found in north-west Europe;
- H3b and H3k, found in the British Isles and Catalonia;
- H3c, found in Western Europe, including among the Basques;
- H3h, found throughout northern Europe, including the remains of Cerdic (519 to 534), King of Wessex;[32]
- H3i found in Ireland and Scotland;
- H3j found in Italy;
- H3v found especially in Germanic countries and;
- H3z found in Atlantic Europe.
The basal H3* haplogroup is found among the Tuareg inhabiting the Gossi area in Mali (4.76%).[24]
H5
H5 may have evolved in West Asia, where it is most frequent and diverse in the Western Caucasus. However, its H5a subclade has a stronger representation in Europe, though at low levels.[33]
H2, H6 and H8
The H2, H6 and H8 haplogroups are somewhat common in Eastern Europe and the Caucasus.[21] They may be the most common H subclades among Central Asians and have also been found in West Asia.[22] H2a5 has been found in the Basque Country, Spain,[34] and in Norway, Ireland and Slovakia.[3] H6a1a1a is common among Ashkenazi Jews.[35]
H4, H7 and H13
These H4, H7 and H13 subhaplogroups are present in both Europe and West Asia; the H13 subclade is also found in the Caucasus; H13c was found in a 9,700 year old sample in Mesolithic Georgia.[36] They are quite rare.[21] H4 is often found in the Iberian peninsula,[34] Britain and Ireland.
H4 and H13, along with H2 account for 42% of the hg H lineages in Egypt.[37]
H10
Haplogroup H10 is subclade which came into existence between 6,300 and 10,900 years ago. Its descendant branches are H10a H10b H10c H10d H10e H10f H10g and H10h.[38]
Haplogroup H10e has been found at a neolithic site, namely the Bom Santo cave near Lisbon. This is the oldest sample of H10 which has ever been found and it has been dated to 3735 BCE (+- 45 years).[39]
H11
H11 is commonly found in Central Europe.[34]
H18
H18 occurs on the Arabian Peninsula. [40]
H20 and H21
These haplogroups are both found in the Caucasus region.[33] H20 also appears at low levels in the Iberian Peninsula (less than 1%), Arabian Peninsula (1%) and Near East (2%).[40]
H22 through H95a
These subclades are found mostly in Europe, South-West Asia and Central Asia.
For example:H53.
H53*, H53a, H53a1, H53b, H53c in Yfull. (found in Basques "probably a basque marker", Irish, Germans, Belarussian, Polish, Amazigh (found in Algeria by Zheng), Canary Islands (found in ancient guanches by Rosa Fregel) probably related with the "Iberomaurusian culture" in North Africa and the ancestors of the Amazigh people.
Also related with the Xinjiang (found in Uyghurs by Zheng), and probably is related with the "Early Agriculture expansion in Central Asia, and far away".
Tree
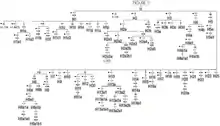
This phylogenetic tree of haplogroup H subclades is based on Build 16(February 2014) of the Phylotree, an internationally accepted standard.[41] The full tree can be viewed at Phylotree.
| mtDNA HG "H" p-tree |
|---|
Also is related with the Xinjiang(found in Uyghurs by Zheng), and probably is related with the "Early Agriculture expansion in Central Asia, and far away".
|
Genetic traits
Haplogroup H was found as a possible increased risk factor for ischemic cardiomyopathy development.[42]
Popular culture
In his popular book The Seven Daughters of Eve, Bryan Sykes named the originator of this mtDNA haplogroup Helena. Stephen Oppenheimer uses the very similar name Helina in his book The Origins of the British.
See also
| Wikimedia Commons has media related to Haplogroup H (mtDNA). |
|
Phylogenetic tree of human mitochondrial DNA (mtDNA) haplogroups | |||||||||||||||||||||||||||||||||||||||
| Mitochondrial Eve (L) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | O | A | S | R | I | W | X | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| H | V | J | T | ||||||||||||||||||||||||||||||||||||
References
- Achilli A, Rengo C, Magri C, Battaglia V, Olivieri A, Scozzari R, et al. (November 2004). "The molecular dissection of mtDNA haplogroup H confirms that the Franco-Cantabrian glacial refuge was a major source for the European gene pool". American Journal of Human Genetics. 75 (5): 910–8. doi:10.1086/425590. PMC 1182122. PMID 15382008.
- https://web.archive.org/web/20190512023944/https://www.yfull.com/mtree/H135/
- van Oven M, Kayser M (February 2009). "Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation". Human Mutation. 30 (2): E386–94. doi:10.1002/humu.20921. PMID 18853457. S2CID 27566749.
- Behar DM, van Oven M, Rosset S, Metspalu M, Loogväli EL, Silva NM, et al. (April 2012). "A "Copernican" reassessment of the human mitochondrial DNA tree from its root". American Journal of Human Genetics. 90 (4): 675–84. doi:10.1016/j.ajhg.2012.03.002. PMC 3322232. PMID 22482806.
- Caramelli D, Milani L, Vai S, Modi A, Pecchioli E, Girardi M, et al. (July 2008). "A 28,000 years old Cro-Magnon mtDNA sequence differs from all potentially contaminating modern sequences". PLOS ONE. 3 (7): e2700. Bibcode:2008PLoSO...3.2700C. doi:10.1371/journal.pone.0002700. PMC 2444030. PMID 18628960.
- Secher B, Fregel R, Larruga JM, Cabrera VM, Endicott P, Pestano JJ, et al. (May 2014). "The history of the North African mitochondrial DNA haplogroup U6 gene flow into the African, Eurasian and American continents". BMC Evolutionary Biology. 14: 109. doi:10.1186/1471-2148-14-109. PMC 4062890. PMID 24885141.
- Kefi R, Hechmi M, Naouali C, Jmel H, Hsouna S, Bouzaid E, et al. (January 2018). "On the origin of Iberomaurusians: new data based on ancient mitochondrial DNA and phylogenetic analysis of Afalou and Taforalt populations". Mitochondrial DNA. Part A, DNA Mapping, Sequencing, and Analysis. 29 (1): 147–157. doi:10.1080/24701394.2016.1258406. PMID 28034339. S2CID 4490910.
- Brotherton P, Haak W, Templeton J, Brandt G, Soubrier J, Jane Adler C, et al. (2013). "Neolithic mitochondrial haplogroup H genomes and the genetic origins of Europeans". Nature Communications. 4: 1764. Bibcode:2013NatCo...4.1764.. doi:10.1038/ncomms2656. PMC 3978205. PMID 23612305.
- Nikitin AG, Potekhina I, Rohland N, Mallick S, Reich D, Lillie M (2017). "Mitochondrial DNA analysis of eneolithic trypillians from Ukraine reveals neolithic farming genetic roots". PLOS ONE. 12 (2): e0172952. Bibcode:2017PLoSO..1272952N. doi:10.1371/journal.pone.0172952. PMC 5325568. PMID 28235025.
- Schuenemann VJ, Peltzer A, Welte B, van Pelt WP, Molak M, Wang CC, et al. (May 2017). "Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods". Nature Communications. 8: 15694. Bibcode:2017NatCo...815694S. doi:10.1038/ncomms15694. PMC 5459999. PMID 28556824.
- Sirak K, Frenandes D, Novak M, Van Gerven D, Pinhasi R (2016). "Abstract Book of the IUAES Inter-Congress 2016 – A community divided? Revealing the community genome(s) of Medieval Kulubnarti using next- generation sequencing". Abstract Book of the Iuaes Inter-Congress 2016. IUAES: 115.
- Ghezzi D, Marelli C, Achilli A, Goldwurm S, Pezzoli G, Barone P, et al. (June 2005). "Mitochondrial DNA haplogroup K is associated with a lower risk of Parkinson's disease in Italians". European Journal of Human Genetics. 13 (6): 748–52. doi:10.1038/sj.ejhg.5201425. PMID 15827561.
- Sykes B (2001). The Seven Daughters of Eve. London; New York: Bantam Press. ISBN 978-0393020182.
- "Maternal Ancestry". Oxford Ancestors. Archived from the original on 15 July 2017. Retrieved 7 February 2013.
- "Haplogroup H". Atlas of the Human Journey – The Genographic Project. National Geographic.
- Metspalu M, Kivisild T, Metspalu E, Parik J, Hudjashov G, Kaldma K, et al. (August 2004). "Most of the extant mtDNA boundaries in south and southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans". BMC Genetics. 5: 26. doi:10.1186/1471-2156-5-26. PMC 516768. PMID 15339343.
- Non A. "Analyses if Genetic Data Within A=an Interdisciplinary Framework to Investigate Recent Human Evolutionary History and Complex Disease" (PDF). University of Florida. Retrieved 3 May 2016.
- Kujanová M, Pereira L, Fernandes V, Pereira JB, Cerný V (October 2009). "Near eastern neolithic genetic input in a small oasis of the Egyptian Western Desert". American Journal of Physical Anthropology. 140 (2): 336–46. doi:10.1002/ajpa.21078. PMID 19425100.
- Stevanovitch A, Gilles A, Bouzaid E, Kefi R, Paris F, Gayraud RP, et al. (January 2004). "Mitochondrial DNA sequence diversity in a sedentary population from Egypt". Annals of Human Genetics. 68 (Pt 1): 23–39. doi:10.1046/j.1529-8817.2003.00057.x. PMID 14748828. S2CID 44901197.
- Cerný V, Pereira L, Kujanová M, Vasíková A, Hájek M, Morris M, Mulligan CJ (April 2009). "Out of Arabia-the settlement of island Soqotra as revealed by mitochondrial and Y chromosome genetic diversity". American Journal of Physical Anthropology. 138 (4): 439–47. doi:10.1002/ajpa.20960. PMID 19012329.
- Pereira L, Richards M, Goios A, Alonso A, Albarrán C, Garcia O, et al. (January 2005). "High-resolution mtDNA evidence for the late-glacial resettlement of Europe from an Iberian refugium". Genome Research. 15 (1): 19–24. doi:10.1101/gr.3182305. PMC 540273. PMID 15632086.
- Loogväli EL, Roostalu U, Malyarchuk BA, Derenko MV, Kivisild T, Metspalu E, et al. (November 2004). "Disuniting uniformity: a pied cladistic canvas of mtDNA haplogroup H in Eurasia". Molecular Biology and Evolution. 21 (11): 2012–21. doi:10.1093/molbev/msh209. PMID 15254257.
- Ottoni C, Primativo G, Hooshiar Kashani B, Achilli A, Martínez-Labarga C, Biondi G, et al. (October 2010). "Mitochondrial haplogroup H1 in north Africa: an early holocene arrival from Iberia". PLOS ONE. 5 (10): e13378. Bibcode:2010PLoSO...513378O. doi:10.1371/journal.pone.0013378. PMC 2958834. PMID 20975840.
- Pereira L, Cerný V, Cerezo M, Silva NM, Hájek M, Vasíková A, et al. (August 2010). "Linking the sub-Saharan and West Eurasian gene pools: maternal and paternal heritage of the Tuareg nomads from the African Sahel". European Journal of Human Genetics. 18 (8): 915–23. doi:10.1038/ejhg.2010.21. PMC 2987384. PMID 20234393.
- Kulichová I, Fernandes V, Deme A, Nováčková J, Stenzl V, Novelletto A, et al. (October 2017). "Internal diversification of non-Sub-Saharan haplogroups in Sahelian populations and the spread of pastoralism beyond the Sahara". American Journal of Physical Anthropology. 164 (2): 424–434. doi:10.1002/ajpa.23285. PMID 28736914.
- Lipson M, Szécsényi-Nagy A, Mallick S, Pósa A, Stégmár B, Keerl V, et al. (November 2017). "Parallel palaeogenomic transects reveal complex genetic history of early European farmers". Nature. 551 (7680): 368–372. Bibcode:2017Natur.551..368L. doi:10.1038/nature24476. PMC 5973800. PMID 29144465.
- Rodríguez-Varela R, Günther T, Krzewińska M, Storå J, Gillingwater TH, MacCallum M, et al. (November 2017). "Genomic Analyses of Pre-European Conquest Human Remains from the Canary Islands Reveal Close Affinity to Modern North Africans". Current Biology. 27 (21): 3396–3402.e5. doi:10.1016/j.cub.2017.09.059. PMID 29107554.
- Alejandra C, Fregel R, Trujillo-Mederos A, Hervella M, De-La-Rúa C, Arnay-De-La-Rosa M (2017). "Genetic studies on the prehispanic population buried in Punta Azul cave (El Hierro, Canary Islands)". Journal of Archaeological Science. 78: 20–28. doi:10.1016/j.jas.2016.11.004.
- "Origins, spread and ethnic association of European haplogroups and subclades". Euopedia.
- Hay M. "Haplogroup H (mtDNA)". Access date 2015/10/02
- Hendrickson SL, Hutcheson HB, Ruiz-Pesini E, Poole JC, Lautenberger J, Sezgin E, et al. (November 2008). "Mitochondrial DNA haplogroups influence AIDS progression". AIDS. 22 (18): 2429–39. doi:10.1097/QAD.0b013e32831940bb. PMC 2699618. PMID 19005266.
- "The Haplogroup H&HV mtGenome Project: H3". mtDNA Test Results for Members. Retrieved 2 October 2015.
- Roostalu U, Kutuev I, Loogväli EL, Metspalu E, Tambets K, Reidla M, et al. (February 2007). "Origin and expansion of haplogroup H, the dominant human mitochondrial DNA lineage in West Eurasia: the Near Eastern and Caucasian perspective". Molecular Biology and Evolution. 24 (2): 436–48. doi:10.1093/molbev/msl173. PMID 17099056.
- Alvarez-Iglesias V, Mosquera-Miguel A, Cerezo M, Quintáns B, Zarrabeitia MT, Cuscó I, et al. (2009). "New population and phylogenetic features of the internal variation within mitochondrial DNA macro-haplogroup R0". PLOS ONE. 4 (4): e5112. Bibcode:2009PLoSO...4.5112A. doi:10.1371/journal.pone.0005112. PMC 2660437. PMID 19340307.
- Costa MD, Pereira JB, Pala M, Fernandes V, Olivieri A, Achilli A, Perego UA, Rychkov S, Naumova O, Hatina J, Woodward SR, Eng KK, Macaulay V, Carr M, Soares P, Pereira L, Richards MB (2013). "A substantial prehistoric European ancestry amongst Ashkenazi maternal lineages". Nature Communications. 4: 2543. Bibcode:2013NatCo...4.2543C. doi:10.1038/ncomms3543. PMC 3806353. PMID 24104924.
- Jones, Eppie R.; Gonzalez-Fortes, Gloria; Connell, Sarah; Siska, Veronika; Eriksson, Anders; Martiniano, Rui; McLaughlin, Russell L.; Gallego Llorente, Marcos; Cassidy, Lara M.; Gamba, Cristina; Meshveliani, Tengiz; Bar-Yosef, Ofer; Müller, Werner; Belfer-Cohen, Anna; Matskevich, Zinovi; Jakeli, Nino; Higham, Thomas F. G.; Currat, Mathias; Lordkipanidze, David; Hofreiter, Michael; Manica, Andrea; Pinhasi, Ron; Bradley, Daniel G. (16 November 2015). "Upper Palaeolithic genomes reveal deep roots of modern Eurasians". Nature Communications. 6 (1): 8912. Bibcode:2015NatCo...6.8912J. doi:10.1038/ncomms9912. PMC 4660371. PMID 26567969.
- Bekada A, Fregel R, Cabrera VM, Larruga JM, Pestano J, Benhamamouch S, González AM (2013). "Introducing the Algerian mitochondrial DNA and Y-chromosome profiles into the North African landscape". PLOS ONE. 8 (2): e56775. Bibcode:2013PLoSO...856775B. doi:10.1371/journal.pone.0056775. PMC 3576335. PMID 23431392.
- Behar DM, van Oven M, Rosset S, Metspalu M, Loogväli EL, Silva NM, Kivisild T, Torroni A, Villems R (April 2012). "A "Copernican" reassessment of the human mitochondrial DNA tree from its root". American Journal of Human Genetics. 90 (4): 675–84. doi:10.1016/j.ajhg.2012.03.002. PMC 3322232. PMID 22482806.
- Faustino de Carvalho A (2014). Bom Santo cave (Lisbon) and the middle neolithic societies of southern Portugal. Faro: Universidade do Algarvede. ISBN 9789899766631. OCLC 946308166.
- Ennafaa H, Cabrera VM, Abu-Amero KK, González AM, Amor MB, Bouhaha R, et al. (February 2009). "Mitochondrial DNA haplogroup H structure in North Africa". BMC Genetics. 10: 8. doi:10.1186/1471-2156-10-8. PMC 2657161. PMID 19243582.
- "PhyloTree.org mtDNA subtree R0". Archived from the original on 2015-12-18.
- Fernández-Caggiano M, Barallobre-Barreiro J, Rego-Pérez I, Crespo-Leiro MG, Paniagua MJ, Grillé Z, et al. (2012). "Mitochondrial haplogroups H and J: risk and protective factors for ischemic cardiomyopathy". PLOS ONE. 7 (8): e44128. Bibcode:2012PLoSO...744128F. doi:10.1371/journal.pone.0044128. PMC 3429437. PMID 22937160.
External links
- General
- Ian Logan's Mitochondrial DNA Site
- Mannis van Oven's Phylotree
- Haplogroup H
- mtDNA Haplogroup H Project at Family Tree DNA
- National Geographic's Spread of Haplogroup H, from National Geographic
- mtDNA Haplogroup H article at SNPedia
- Amelia's Helena
- Loogväli EL, Roostalu U, Malyarchuk BA, Derenko MV, Kivisild T, Metspalu E, et al. (November 2004). "Disuniting uniformity: a pied cladistic canvas of mtDNA haplogroup H in Eurasia". Molecular Biology and Evolution. 21 (11): 2012–21. doi:10.1093/molbev/msh209. PMID 15254257.
- Genebase's Tutorials on mtDNA Haplogroup H
- Genebase's Phylogenetic tree of mtDNA Haplogroup H
- Genebase's Geographical distribution of mtDNA Haplogroup H
- Haplogroup and Subcluster Frequencies for European Populations: Helgason A, Hickey E, Goodacre S, Bosnes V, Stefánsson K, Ward R, et al. (March 2001). "mtDna and the islands of the North Atlantic: estimating the proportions of Norse and Gaelic ancestry". American Journal of Human Genetics. 68 (3): 723–37. doi:10.1086/318785. PMC 1274484. PMID 11179019.
- Danish Demes Regional DNA Project: mtDNA Haplogroup H
- Survey/study open to men who are in any branch of Haplogroup H
- Survey/study open to women who are in any branch of Haplogroup H
- Haplogroup H1
- Hope The H1 mtDNA Haplogroup Project