Microbiome
The word microbiome (from the Greek micro meaning "small" and bíos meaning "life") was first used by J.L. Mohr in 1952 in The Scientific Monthly to mean the microorganisms found in a specific environment.[2][3] It was defined in 1988 by Whipps et al. as "a characteristic microbial community occupying a reasonably well-defined habitat which has distinct physio-chemical properties. The term thus not only refers to the microorganisms involved but also encompasses their theatre of activity".[4]
In 2020, an international panel of experts published the outcome of their discussions on the definition of the microbiome.[1] They proposed a definition of the microbiome based on a revival of the "compact, clear, and comprehensive description of the term" as originally provided by Whipps et al., but supplemented with two explanatory sentences.[1]
The first explanatory sentence pronounces the dynamic character of the microbiome:
- The microbiome is defined as a characteristic microbial community occupying a reasonably well-defined habitat which has distinct physio-chemical properties. The microbiome not only refers to the microorganisms involved but also encompass their theatre of activity, which results in the formation of specific ecological niches. The microbiome, which forms a dynamic and interactive micro-ecosystem prone to change in time and scale, is integrated in macro-ecosystems including eukaryotic hosts, and here crucial for their functioning and health.[1]
The second explanatory sentence clearly separates the term microbiota from the term microbiome:
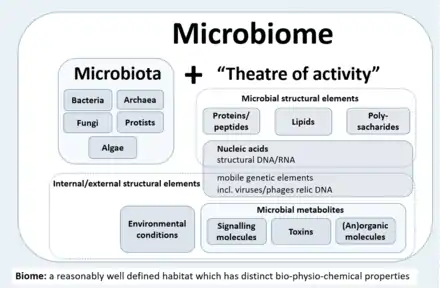
- The microbiota consists of the assembly of microorganisms belonging to different kingdoms (Prokaryotes [Bacteria, Archaea], Eukaryotes [e.g., Protozoa, Fungi, and Algae]), while their theatre of activity includes microbial structures, metabolites, mobile genetic elements (such as transposons, phages, and viruses), and relic DNA embedded in the environmental conditions of the habitat.[1]
Background

Microbiome research originated in microbiology and started back in the seventeenth century. The development of new techniques and equipment has boosted microbiological research and caused paradigm shifts in understanding health and disease. Since infectious diseases have affected human populations throughout most of history, medical microbiology was the earliest focus of research and public interest. Additionally, food microbiology is an old field of empirical applications. The development of the first microscopes allowed the discovery of a new, unknown world and led to the identification of microorganisms.[1]
Access to the previously invisible world opened the eyes and the minds of the researchers of the seventeenth century. Antonie van Leeuwenhoek investigated diverse bacteria of various shapes, fungi, and protozoa, which he called animalcules, mainly from water, mud, and dental plaque samples, and discovered biofilms as a first indication of microorganisms interacting within complex communities. Robert Koch's explanation of the origin of human and animal diseases as a consequence of microbial infection and development of the concept of pathogenicity was an important milestone in microbiology. These findings shifted the focus of the research community and the public on the role of microorganisms as disease-forming agents that needed to be eliminated.[1]
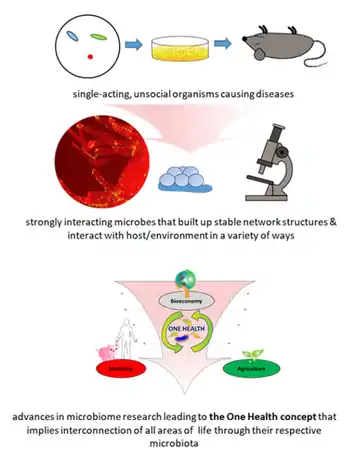
However, comprehensive research over the past century has shown only a small proportion of microorganisms are associated with disease or pathogenicity. The overwhelming majority of microbes are essential for ecosystem functioning and known for beneficial interactions with other microbes as well as macroorganisms. At the end of the nineteenth century, microbial ecology started with the pioneering work by Martinus W. Beijerinck and Sergei Winogradsky. The newly established science of environmental microbiology resulted in another paradigm shift: microorganisms are everywhere in natural environments, often associated with hosts and, for the first time, beneficial effects on their hosts were reported.[5][6][1]
Subsequently, the concept that microorganisms exist as single cells began to change as it became increasingly obvious that microbes occur within complex assemblages in which species interactions and communication are critical to population dynamics and functional activities.[7] Discovery of DNA, the development of sequencing technologies, PCR, and cloning techniques enabled the investigation of microbial communities using cultivation-independent, DNA and RNA-based approaches.[8][1]
A further important step was the introduction of phylogenetic markers such as the 16S rRNA gene for microbial community analysis by Carl Woese and George E. Fox in 1977.[9] Today, we are able to barcode bacteria, archaea, fungi, algae, and protists in their natural habitats, e.g., by targeting their 16S and 18S rRNA genes, internal transcribed spacer (ITS), or, alternatively, specific functional regions of genes coding for specific enzymes.[10][11][12][1]
Another major paradigm shift was initiated at the beginning of this century and continues through today, as new sequencing technologies and accumulated sequence data have highlighted both the ubiquity of microbial communities in association within higher organisms and the critical roles of microbes in human, animal, and plant health.[13] These new possibilities have revolutionized microbial ecology, because the analysis of genomes and metagenomes in a high-throughput manner provides efficient methods for addressing the functional potential of individual microorganisms as well as of whole communities in their natural habitats.[14][15] Multiomics technologies including metatranscriptome, metaproteome and metabolome approaches now provide detailed information on microbial activities in the environment. Based on the rich foundation of data, the cultivation of microbes, which was often ignored or underestimated over the last thirty years, has gained new importance, and high throughput culturomics is now an important part of the toolbox to study microbiomes. The high potential and power of combining multiple "omics" techniques to analyze host-microbe interactions are highlighted in several reviews.[16][17][1]
Defining the microbiome
Microbial communities have commonly been defined as the collection of microorganisms living together. More specifically, microbial communities are defined as multi-species assemblages, in which (micro) organisms interact with each other in a contiguous environment.[18] In 1988, Whipps and colleagues working on the ecology of rhizosphere microorganisms provided the first definition of the term microbiome.[4] They described the microbiome as a combination of the words micro and biome, naming a "characteristic microbial community" in a "reasonably well-defined habitat which has distinct physio-chemical properties" as their "theatre of activity". This definition represents a substantial advancement of the definition of a microbial community, as it defines a microbial community with distinct properties and functions and its interactions with its environment, resulting in the formation of specific ecological niches.[1]
However, many other microbiome definitions have been published in the last few decades. The currently most cited definition by Lederberg[19] describes microbiomes within an ecological context, as a community of commensal, symbiotic, and pathogenic microorganisms within a body space or other environment. Marchesi and Ravel focused in their definition on the genomes and microbial (and viral) gene expression patterns and proteomes in a given environment and its prevailing biotic and abiotic conditions.[20] All these definitions imply that general concepts of macro-ecology could be easily applied to microbe-microbe as well as to microbe-host interactions. However, the extent to which these concepts, developed for macro-eukaryotes, can be applied to prokaryotes with their different lifestyles regarding dormancy, variation of phenotype, and horizontal gene transfer[21] as well as to micro-eukaryotes that is not quite clear. This raises the challenge of considering an entirely novel body of conceptual ecology models and theory for microbiome ecology, particularly in relation to the diverse hierarchies of interactions of microbes with one another and with the host biotic and abiotic environments. Many current definitions fail to capture this complexity and describe the term microbiome as encompassing the genomes of microorganisms only (see table ↓).[1]
| Microbiome definitions[1] | |
|---|---|
| Definition type | Examples |
| Ecological | Definitions based on ecology describe the microbiome following the concepts derived from the ecology of multicellular organisms. The main issue here is that the theories from the macro-ecology do not always fit the rules in the microbial world. |
| |
| Organisms/host-dependent | The host-dependent definitions are based on the microbial interactions with the host. The main gaps here concern the question whether the microbial-host interaction data gained from one host can be transferred to another. The understanding of coevolution and selection in the host-dependent definitions is also underrepresented. |
| |
| Genomic/ method-driven | There is a variety of microbiome definitions available that are driven by the methods applied. Mostly, these definitions rely on DNA sequence-based analysis and describe microbiome as a collective genome of microorganisms in a specific environment. The main bottleneck here is that every new available technology will result in a need for a new definition. |
| |
| Combined | There are some microbiome definitions available that fit several categories with their advantages and disadvantages. |
| |
In 2020, a panel of international experts, organised by the EU-funded MicrobiomeSupport project,[32] published the results of their deliberations on the definition of the microbiome.[1] The panel was composed of about 40 leaders from diverse microbiome areas, and about one hundred further experts from around the world contributed through an online survey. They proposed a definition of the microbiome based on a revival of the compact, clear, and comprehensive description of the term as originally provided by Whipps et al. in 1988,[4] amended with a set of recommendations considering subsequent technological developments and research findings. They clearly separate the terms microbiome and microbiota and provide a comprehensive discussion considering the composition of microbiota, the heterogeneity and dynamics of microbiomes in time and space, the stability and resilience of microbial networks, the definition of core microbiomes, and functionally relevant keystone species as well as co-evolutionary principles of microbe-host and inter-species interactions within the microbiome.[1]
The panel extended the Whipps et al. definition, which contains all important points that are valid even 30 years after its publication in 1988, by two explanatory sentences differentiating the terms microbiome and microbiota and pronouncing its dynamic character, as follows:
- The microbiome is defined as a characteristic microbial community occupying a reasonable well-defined habitat which has distinct physio-chemical properties. The microbiome not only refers to the microorganisms involved but also encompass their theatre of activity, which results in the formation of specific ecological niches. The microbiome, which forms a dynamic and interactive micro-ecosystem prone to change in time and scale, is integrated in macro-ecosystems including eukaryotic hosts, and here crucial for their functioning and health.[1]
- The microbiota consists of the assembly of microorganisms belonging to different kingdoms (prokaryotes (bacteria, archaea), eukaryotes (algae, protozoa, fungi etc), while "their theatre of activity" includes microbial structures, metabolites, mobile genetic elements (such as transposons, phages, and viruses), and relic DNA embedded in the environmental conditions of the habitat.[1]
Microbiota – members of the microbiome
The microbiota comprises all living members forming the microbiome. Most microbiome researchers agree bacteria, archaea, fungi, algae, and small protists should be considered as members of the microbiome.[20][1] The integration of phages, viruses, plasmids, and mobile genetic elements is a more controversial issue in the definition of the microbiome. There is also no clear consensus as to whether extracellular DNA derived from dead cells, so-called "relic DNA", belongs to the microbiome.[33][1] Relic DNA can be up to 40% of the sequenced DNA in soil,[34] and was up to 33% of the total bacterial DNA on average in a broader analysis of habitats with the highest proportion of 80% in some samples.[35] Despite its omnipresence and abundance, relic DNA had a minimal effect on estimates of taxonomic and phylogenetic diversity.[35][1]
When it comes to the use of specific terms, a clear differentiation between microbiome and microbiota helps to avoid the controversy concerning the members of a microbiome.[1] Microbiota is usually defined as the assemblage of living microorganisms present in a defined environment.[20] As phages, viruses, plasmids, prions, viroids, and free DNA are usually not considered as living microorganisms,[36] they do not belong to the microbiota.[1]
The term microbiome, as it was originally postulated by Whipps and coworkers,[4] includes not only the community of the microorganisms but also their "theatre of activity". The latter involves the whole spectrum of molecules produced by the microorganisms, including their structural elements (nucleic acids, proteins, lipids, polysaccharides), metabolites (signalling molecules, toxins, organic, and inorganic molecules), and molecules produced by coexisting hosts and structured by the surrounding environmental conditions. Therefore, all mobile genetic elements, such as phages, viruses, and "relic" and extracellular DNA, should be included in the term microbiome, but are not a part of microbiota. The term microbiome is also sometimes confused with the metagenome. Metagenome is, however, clearly defined as a collection of genomes and genes from the members of a microbiota.[20][1]
Microbiome studies sometimes focus on the behaviour of a specific group of microbiota, generally in relation to or justified by a clear hypothesis. More and more terms like bacteriome, archaeome, mycobiome, or virome have started appearing in the scientific literature, but these terms do not refer to biomes (a regional ecosystem with a distinct assemblage of (micro) organisms, and physical environment often reflecting a certain climate and soil) as the microbiome itself.[1] Consequently, it would be better to use the original terms (bacterial, archaeal, or fungal community). In contrast to the microbiota, which can be studied separately, the microbiome is always composed by all members, which interact with each other, live in the same habitat, and form their ecological niche together. The well-established term virome is derived from virus and genome and is used to describe viral shotgun metagenomes consisting of a collection of nucleic acids associated with a particular ecosystem or holobiont.[37] Viral metagenomes can be suggested as a semantically and scientifically better term.[1]
Plant microbiomes
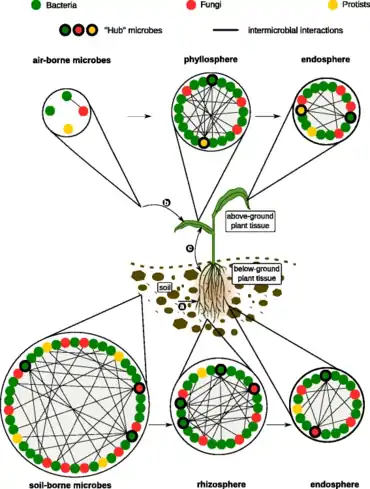
The diagram on the right →
illustrates microbial communities in the soil, air, rhizosphere, phyllosphere, and inside plant tissue (endosphere). In each of these habitats, microbes (represented by colored circles) could interact positively, negatively, or do not interact with other microbes (no lines). Specific microbes, often defined as “hub” or “keystone” species (circles highlighted in bold), are highly connected to other microbes within the networks and likely exert a stronger influence on the structure of microbial communities. (a) Root-associated microbes mainly derive from the soil biome. (b) Leaf-associated microbes originate from various sources such as aerosols, insects, or dust. (c) Relocation between aboveground and belowground microbiota members.[38]
The microbial component of healthy seeds – the seed microbiome – appears to be inherited between plant generations and can dynamically influence germination, plant performance, and survival.[39] As such, methods to optimize the seed microbiomes of major crops could have far-reaching implications for plant breeding and crop improvement to enhance agricultural food, feed, and fiber production.[40]
Marine microbiomes
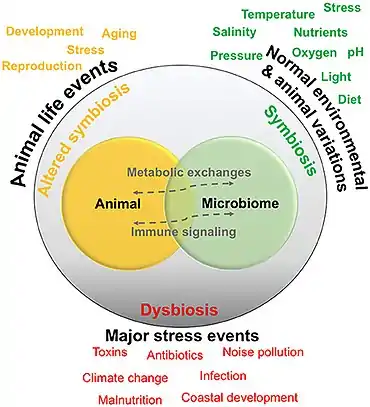
All animals on Earth form associations with microorganisms, including protists, bacteria, archaea, fungi, and viruses. In the ocean, animal–microbial relationships were historically explored in single host–symbiont systems. However, new explorations into the diversity of microorganisms associating with diverse marine animal hosts is moving the field into studies that address interactions between the animal host and a more multi-member microbiome. The potential for microbiomes to influence the health, physiology, behavior, and ecology of marine animals could alter current understandings of how marine animals adapt to change, and especially the growing climate-related and anthropogenic-induced changes already impacting the ocean environment.[41]
The microbiomes of diverse marine animals are currently under study, from simplistic organisms including sponges[42] and ctenophores [43] to more complex organisms such as sea squirts[44] and sharks.[45][41]
The relationship between the Hawaiian bobtail squid and the bioluminescent bacterium Aliivibrio fischeri is one of the best studied symbiotic relationships in the sea and is a choice system for general symbiosis research. This relationship has provided insight into fundamental processes in animal-microbial symbioses, and especially biochemical interactions and signaling between the host and bacterium.[46][47][41]
The gutless marine oligochaete worm Olavius algarvensis is another relatively well-studied marine host to microbes. These three centimetre long worms reside within shallow marine sediments of the Mediterranean Sea. The worms do not contain a mouth or a digestive or excretory system, but are instead nourished with the help of a suite of extracellular bacterial endosymbionts that reside upon coordinated use of sulfur present in the environment.[48] This system has benefited from some of the most sophisticated 'omics and visualization tools.[49] For example, multi-labeled probing has improved visualization of the microbiome[50] and transcriptomics and proteomics have been applied to examine host–microbiome interactions, including energy transfer between the host and microbes[51] and recognition of the consortia by the worm's innate immune system.[52] The major strength of this system is that it does offer the ability to study host–microbiome interactions with a low diversity microbial consortium, and it also offers a number of host and microbial genomic resources[49][53][41]
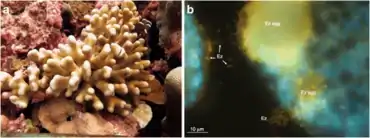
Corals are one of the more common examples of an animal host whose symbiosis with microalgae can turn to dysbiosis, and is visibly detected as bleaching. Coral microbiomes have been examined in a variety of studies, which demonstrate how variations in the ocean environment, most notably temperature, light, and inorganic nutrients, affect the abundance and performance of the microalgal symbionts, as well as calcification and physiology of the host.[55][56] Studies have also suggested that resident bacteria, archaea, and fungi additionally contribute to nutrient and organic matter cycling within the coral, with viruses also possibly playing a role in structuring the composition of these members, thus providing one of the first glimpses at a multi-domain marine animal symbiosis.[57] The gammaproteobacterium Endozoicomonas is emerging as a central member of the coral's microbiome, with flexibility in its lifestyle.[54][58] Given the recent mass bleaching occurring on reefs,[59] corals will likely continue to be a useful and popular system for symbiosis and dysbiosis research.[41]
Sponges are common members of the ocean's diverse benthic habitats and their abundance and ability to filter large volumes of seawater have led to the awareness that these organisms play critical roles in influencing benthic and pelagic processes in the ocean.[60] They are one of the oldest lineages of animals, and have a relatively simple body plan that commonly associates with bacteria, archaea, algal protists, fungi, and viruses.[61] Sponge microbiomes are composed of specialists and generalists, and complexity of their microbiome appears to be shaped by host phylogeny.[62] Studies have shown that the sponge microbiome contributes to nitrogen cycling in the oceans, especially through the oxidation of ammonia by archaea and bacteria.[63][64] Most recently, microbial symbionts of tropical sponges were shown to produce and store polyphosphate granules,[65] perhaps enabling the host to survive periods of phosphate depletion in oligotrophic marine environments.[66] The microbiomes of some sponge species do appear to change in community structure in response to changing environmental conditions, including temperature[67] and ocean acidification,[68][69] as well as synergistic impacts.[70]
Underlying complexity
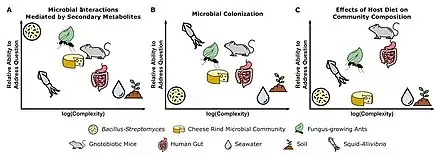
Each microbiome system is suited to address different types of questions based on the culturability of microbes, genetic tractability of microbes and host (where relevant), ability to maintain system in laboratory setting, and ability to make host/environment germfree.[71]
Three different systems are shown in the figure on the right. (A) Pairwise interactions between the soil bacteria Bacillus subtilis and Streptomyces spp. are well-suited for characterizing the functions of secondary metabolites in microbial interactions. (B) The symbiosis between bobtail squid and the marine bacterium Aliivibrio fischeri is fundamental to understanding host and microbial factors that influence colonization. (C) The use of gnotobiotic mice is crucial for making links between host diet and the effects on specific microbial taxa in a community.[71]
Host-microbe coevolution
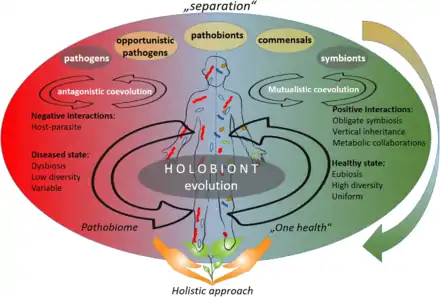
According to the "separation" approach (upper part of the figure on the right), the microorganisms can be divided into pathogens, neutral, and symbionts, depending on their interaction with their host. The coevolution between host and its associated microbiota may be accordingly described as antagonistic (based on negative interactions) or mutualistic (based on positive interactions).[1]
As of 2020, the emergence in publications about opportunistic pathogens and pathobionts has produced a shift towards a holistic approach in the coevolutions theory (lower part of the figure on the right). The holistic approach sees the host and its associated microbiota as one unit (the so-called holobiont), that coevolves as one entity. According to the holistic approach, holobiont's disease state is linked to dysbiosis, low diversity of the associated microbiota, and their variability: a so-called pathobiome state. The healthy state, on the other hand, is accompanied with eubiosis, high diversity, and uniformity of the respective microbiota.[1]
See also
References
- Berg, Gabriele; Daria Rybakova, Doreen Fischer, Tomislav Cernava, Marie-Christine Champomier Vergès, Trevor Charles, Xiaoyulong Chen, Luca Cocolin, Kellye Eversole, Gema Herrero Corral, Maria Kazou, Linda Kinkel, Lene Lange, Nelson Lima, Alexander Loy, James A. Macklin, Emmanuelle Maguin, Tim Mauchline, Ryan McClure, Birgit Mitter, Matthew Ryan, Inga Sarand, Hauke Smidt, Bettina Schelkle, Hugo Roume, G. Seghal Kiran, Joseph Selvin, Rafael Soares Correa de Souza, Leo van Overbeek, Brajesh K. Singh, Michael Wagner, Aaron Walsh, Angela Sessitsch and Michael Schloter (2020) "Microbiome definition re-visited: old concepts and new challenges". Microbiome, 8(103): 1–22. doi:10.1186/s40168-020-00875-0.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License. - "BioConcepts". www.biological-concepts.com. Retrieved 2020-12-18.
- "microbiome". Oxford English Dictionary (Online ed.). Oxford University Press. Retrieved 2020-12-18. (Subscription or participating institution membership required.)
- Whipps J., Lewis K. and Cooke R. (1988) "Mycoparasitism and plant disease control". In: Burge M (Ed.) Fungi in Biological Control Systems, Manchester University Press, pages 161–187. ISBN 9780719019791.
- Hiltner L. (1902) "Die Keimungsverhältnisse der Leguminosensamen und ihre Beeinflussung durch Organismenwirkung". In: Parey P and Springer J (Eds.) Arb Biol Abt Land u Forstw K Gsndhtsamt, 3, Berlin. Pages 1-545.
- Metchnikoff E. The prolongation of life: optimistic studies. GP Putnam's Sons; 1908.
- Bassler, B.L. (2002) "Small talk: cell-to-cell communication in bacteria". Cell, 109(4): 421–424. doi:10.1016/S0092-8674(02)00749-3.
- Brul, S., Kallemeijn, W. and Smits, G. (2008) "Functional genomics for food microbiology: molecular mechanisms of weak organic-acid preservative adaptation in yeast". CAB Rev, 3: 1–14. doi:10.1079/PAVSNNR20083005.
- Woese, C.R. and Fox, G.E. (1977) "Phylogenetic structure of the prokaryotic domain: the primary kingdoms". Proceedings of the National Academy of Sciences, 74(11): 5088–5090. doi:10.1073/pnas.74.11.5088.
- Uksa, M., Schloter, M., Endesfelder, D., Kublik, S., Engel, M., Kautz, T., Köpke, U. and Fischer, D. (2015) "Prokaryotes in subsoil—evidence for a strong spatial separation of different phyla by analysing co-occurrence networks". Frontiers in microbiology, 6: 1269. doi:10.3389/fmicb.2015.01269.
- Maritz, J.M., Rogers, K.H., Rock, T.M., Liu, N., Joseph, S., Land, K.M. and Carlton, J.M. (2017) "An 18S rRNA workflow for characterizing protists in sewage, with a focus on zoonotic trichomonads". Microbial ecology, 74(4): 923–936. doi:10.1007/s00248-017-0996-9.
- Purahong, W., Wubet, T., Lentendu, G., Schloter, M., Pecyna, M.J., Kapturska, D., Hofrichter, M., Krüger, D. and Buscot, F. (2016) "Life in leaf litter: novel insights into community dynamics of bacteria and fungi during litter decomposition". Molecular Ecology, 25(16): 4059–4074. doi:10.1111/mec.13739.
- Lozupone, C.A., Stombaugh, J.I., Gordon, J.I., Jansson, J.K. and Knight, R. (2012) "Diversity, stability and resilience of the human gut microbiota". Nature, 489(7415): 220–230. doi:10.1038/nature11550.
- Venter, J.C., Remington, K., Heidelberg, J.F., Halpern, A.L., Rusch, D., Eisen, J.A., Wu, D., Paulsen, I., Nelson, K.E., Nelson, W. and Fouts, D.E. (2004) "Environmental genome shotgun sequencing of the Sargasso Sea". Science, 304(5667): 66–74. doi:10.1126/science.1093857.
- Liu, L., Li, Y., Li, S., Hu, N., He, Y., Pong, R., Lin, D., Lu, L. and Law, M. (2012) "Comparison of next-generation sequencing systems". BioMed Research International, 2012: 251364. doi:10.1155/2012/251364.
- Stegen, J.C., Bottos, E.M. and Jansson, J.K. (2018) "A unified conceptual framework for prediction and control of microbiomes". Current Opinion in Microbiology, 44: 20–27. doi:10.1016/j.mib.2018.06.002.
- Knight, R., Vrbanac, A., Taylor, B.C., Aksenov, A., Callewaert, C., Debelius, J., Gonzalez, A., Kosciolek, T., McCall, L.I., McDonald, D. and Melnik, A.V. (2018) "Best practices for analysing microbiomes". Nature Reviews Microbiology, 16(7): 410–422. doi:10.1038/s41579-018-0029-9.
- Konopka, A. (2009) "What is microbial community ecology?" The ISME Journal, 3(11): 1223–1230. {{doi|Konopka, A., 2009. What is microbial community ecology?. The ISME journal, 3(11), pp.1223-1230. doi:10.1038/ismej.2009.88.
- Lederberg, J. and McCray, A.T. (2001) "'Ome Sweet'Omics--A genealogical treasury of words". The Scientist, 15(7): 8.
- Marchesi, J.R. and Ravel, J. (2015) "The vocabulary of microbiome research: a proposal". Microbiome, 3(31). doi:10.1186/s40168-015-0094-5.
- Prosser, J.I., Bohannan, B.J., Curtis, T.P., Ellis, R.J., Firestone, M.K., Freckleton, R.P., Green, J.L., Green, L.E., Killham, K., Lennon, J.J. and Osborn, A.M. (2007) "The role of ecological theory in microbial ecology". Nature Reviews Microbiology, 5(5): 384–392. doi:10.1038/nrmicro1643.
- del Carmen Orozco-Mosqueda, M., del Carmen Rocha-Granados, M., Glick, B.R. and Santoyo, G. (2018) "Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms". Microbiological Research, 208: 25–31. doi:10.1016/j.micres.2018.01.005.
- Merriam-Webster Dictionary – microbiome.
- Human Microbiome Project. Accessed 25 Aug 2020.
- Nature.com: Microbiome. Accessed 25 August 2020.
- ScienceDirect: Microbiome Accessed 25 August 2020.
- Arevalo, P., VanInsberghe, D., Elsherbini, J., Gore, J. and Polz, M.F. (2019) "A reverse ecology approach based on a biological definition of microbial populations". Cell, 178(4): 820–834. doi:10.1016/j.cell.2019.06.033.
- Schlaeppi, K. and Bulgarelli, D. (2015) "The plant microbiome at work". Molecular Plant-Microbe Interactions, 28(3): 212–217. doi:10.1094/MPMI-10-14-0334-FI.
- Rogers Y-H and Zhang C. (2016) "Genomic Technologies in Medicine and Health: Past, Present, and Future". In: Kumar D and Antonarakis S. (Eds.) Medical and Health Genomics. Oxford: Academic Press, pages 15–28. ISBN 9780127999227.
- Ho, H.E. and Bunyavanich, S. (2018) "Role of the microbiome in food allergy". Current allergy and asthma reports, 18(4): 27. doi:10.1007/s11882-018-0780-z.
- Whiteside, S.A., Razvi, H., Dave, S., Reid, G. and Burton and J.P. (2015) "The microbiome of the urinary tract—a role beyond infection". Nature Reviews Urology, 12(2): 81–90. doi:10.1038/nrurol.2014.361.
- MicrobiomeSupport project
- Carini, Paul (2016) A census of the dead: the story behind microbial ‘relic DNA’ in soil Nature Research: Microbiology.
- Carini, P., Marsden, P.J., Leff, J.W., Morgan, E.E., Strickland, M.S. and Fierer, N. (2016) "Relic DNA is abundant in soil and obscures estimates of soil microbial diversity". Nature Microbiology, 2(3): 1–6. doi:10.1038/nmicrobiol.2016.242.
- Lennon, J.T., Muscarella, M.E., Placella, S.A. and Lehmkuhl, B.K. (2018) "How, when, and where relic DNA affects microbial diversity". mBio, 9(3). doi:10.1128/mBio.00637-18.
- Dupré JO, O’Malley MA (2009) "Varieties of living things: life at the intersection of lineage and metabolism". In: Normandin S and Wolfe C (Eds.) Vitalism and the Scientific Image in Post-Enlightenment Life Science 1800–2010. Dordrecht: Springer, pages 311–344. ISBN 9789400724457.
- McDaniel, L., Breitbart, M., Mobberley, J., Long, A., Haynes, M., Rohwer, F. and Paul, J.H., 2008. Metagenomic analysis of lysogeny in Tampa Bay: implications for prophage gene expression. PLoS One, 3(9), p.e3263. doi:10.1371/journal.pone.0003263.
- Hassani, M.A., Durán, P. and Hacquard, S. (2018) "Microbial interactions within the plant holobiont". Microbiome, 6(1): 58. doi:10.1186/s40168-018-0445-0. Material was copied from this source, which is available under a [https://creativecommons.org/licenses/by/4.0/ Creative Commons Attribution 4.0 International License
- Matsumoto, Haruna; Fan, Xiaoyan; Wang, Yue; Kusstatscher, Peter; Duan, Jie; Wu, Sanling; Chen, Sunlu; Qiao, Kun; Wang, Yiling; Ma, Bin; Zhu, Guonian (2021). "Bacterial seed endophyte shapes disease resistance in rice". Nature Plants. 7 (1): 60–72. doi:10.1038/s41477-020-00826-5. ISSN 2055-0278.
- Mitter, B., Pfaffenbichler, N., Flavell, R., Compant, S., Antonielli, L., Petric, A., Berninger, T., Naveed, M., Sheibani-Tezerji, R., von Maltzahn, G. and Sessitsch, A. (2017) "A new approach to modify plant microbiomes and traits by introducing beneficial bacteria at flowering into progeny seeds". Frontiers in Microbiology, 8: 11. doi:10.3389/fmicb.2017.00011. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Apprill, A. (2017) "Marine animal microbiomes: toward understanding host–microbiome interactions in a changing ocean". Frontiers in Marine Science, 4: 222. doi:10.3389/fmars.2017.00222. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Webster, N.S., Negri, A.P., Botté, E.S., Laffy, P.W., Flores, F., Noonan, S., Schmidt, C. and Uthicke, S. (2016) "Host-associated coral reef microbes respond to the cumulative pressures of ocean warming and ocean acidification". Scientific reports, 6: 19324. doi:10.1038/srep19324.
- Daniels, C. and Breitbart, M. (2012) "Bacterial communities associated with the ctenophores Mnemiopsis leidyi and Beroe ovata". FEMS Microbiology Ecology, 82(1): 90–101. doi:10.1111/j.1574-6941.2012.01409.x.
- Blasiak, L.C., Zinder, S.H., Buckley, D.H. and Hill, R.T. (2014) "Bacterial diversity associated with the tunic of the model chordate Ciona intestinalis". The ISME Journal, 8(2): 309–320. doi:10.1038/ismej.2013.156.
- Givens, C.E., Ransom, B., Bano, N. and Hollibaugh, J.T. (2015) "Comparison of the gut microbiomes of 12 bony fish and 3 shark species". Marine Ecology Progress Series, 518: 209–223. doi:10.3354/meps11034.
- McFall-Ngai, M.J. (2000) "Negotiations between animals and bacteria: the ‘diplomacy’of the squid-vibrio symbiosis". Comparative Biochemistry and Physiology, Part A: Molecular & Integrative Physiology, 126(4): 471–480. doi:10.1016/S1095-6433(00)00233-6.
- McFall-Ngai, M. (2014) "Divining the essence of symbiosis: insights from the squid-vibrio model". PLoS Biology, 12(2): e1001783. doi:10.1371/journal.pbio.1001783.
- Dubilier, N., Mülders, C., Ferdelman, T., de Beer, D., Pernthaler, A., Klein, M., Wagner, M., Erséus, C., Thiermann, F., Krieger, J. and Giere, O. (2001) "Endosymbiotic sulphate-reducing and sulphide-oxidizing bacteria in an oligochaete worm". Nature, 411(6835): 298–302. doi:10.1038/35077067.
- Woyke, T., Teeling, H., Ivanova, N.N., Huntemann, M., Richter, M., Gloeckner, F.O., Boffelli, D., Anderson, I.J., Barry, K.W., Shapiro, H.J. and Szeto, E. (2006) "Symbiosis insights through metagenomic analysis of a microbial consortium". Nature, 443(7114): 950–955. doi:10.1038/nature05192.
- Schimak, M.P., Kleiner, M., Wetzel, S., Liebeke, M., Dubilier, N. and Fuchs, B.M. (2016) "MiL-FISH: Multilabeled oligonucleotides for fluorescence in situ hybridization improve visualization of bacterial cells". Applied and Environmental Microbiology, 82(1): 62–70. doi:10.1128/AEM.02776-15.
- Kleiner, M., Wentrup, C., Lott, C., Teeling, H., Wetzel, S., Young, J., Chang, Y.J., Shah, M., VerBerkmoes, N.C., Zarzycki, J. and Fuchs, G. (2012) "Metaproteomics of a gutless marine worm and its symbiotic microbial community reveal unusual pathways for carbon and energy use". Proceedings of the National Academy of Sciences, 109(19): E1173–E1182. doi:10.1073/pnas.1121198109.
- Wippler, J., Kleiner, M., Lott, C., Gruhl, A., Abraham, P.E., Giannone, R.J., Young, J.C., Hettich, R.L. and Dubilier, N. (2016) "Transcriptomic and proteomic insights into innate immunity and adaptations to a symbiotic lifestyle in the gutless marine worm Olavius algarvensis". BMC Genomics, 17(1): 942. doi:10.1186/s12864-016-3293-y.
- Ruehland, C., Blazejak, A., Lott, C., Loy, A., Erséus, C. and Dubilier, N. (2008) "Multiple bacterial symbionts in two species of co‐occurring gutless oligochaete worms from Mediterranean sea grass sediments". Environmental microbiology, 10(12): 3404–3416. doi:10.1111/j.1462-2920.2008.01728.x.
- Neave, M.J., Apprill, A., Ferrier-Pagès, C. and Voolstra, C.R. (2016) "Diversity and function of prevalent symbiotic marine bacteria in the genus Endozoicomonas". Applied Microbiology and Biotechnology, 100(19): 8315–8324. doi:10.1007/s00253-016-7777-0.
- Dubinsky, Z. and Jokiel, P.L. (1994) "Ratio of energy and nutrient fluxes regulates symbiosis between zooxanthellae and corals". Pacific Science, 48(3): 313–324.
- Anthony, K.R., Kline, D.I., Diaz-Pulido, G., Dove, S. and Hoegh-Guldberg, O.(2008) "Ocean acidification causes bleaching and productivity loss in coral reef builders". Proceedings of the National Academy of Sciences, 105(45): 17442–17446. doi:10.1073/pnas.0804478105.
- Bourne, D.G., Morrow, K.M. and Webster, N.S. (2016) "Insights into the coral microbiome: underpinning the health and resilience of reef ecosystems". Annual Review of Microbiology, 70: 317–340. doi:10.1146/annurev-micro-102215-095440.
- Neave, M.J., Michell, C.T., Apprill, A. and Voolstra, C.R. (2017) "Endozoicomonas genomes reveal functional adaptation and plasticity in bacterial strains symbiotically associated with diverse marine hosts". Scientific Reports, 7: 40579. doi:10.1038/srep40579.
- Hughes, T.P., Kerry, J.T., Álvarez-Noriega, M., Álvarez-Romero, J.G., Anderson, K.D., Baird, A.H., Babcock, R.C., Beger, M., Bellwood, D.R., Berkelmans, R. and Bridge, T.C. (2017) "Global warming and recurrent mass bleaching of corals". Nature, 543(7645): 373–377. doi:10.1038/nature21707.
- Bell, J.J. (2008) "The functional roles of marine sponges". Estuarine, Coastal and Shelf Science, 79(3): 341–353. doi:10.1016/j.ecss.2008.05.002.
- Webster, N.S. and Thomas, T. (2016) "The sponge hologenome". mBio, 7(2). doi:10.1128/mBio.00135-16.
- Thomas, T., Moitinho-Silva, L., Lurgi, M., Björk, J.R., Easson, C., Astudillo-García, C., Olson, J.B., Erwin, P.M., López-Legentil, S., Luter, H. and Chaves-Fonnegra, A. (2016) "Diversity, structure and convergent evolution of the global sponge microbiome". Nature Communications, 7(1): 1-12. doi:10.1038/ncomms11870.
- Bayer, K., Schmitt, S. and Hentschel, U. (2008) "Physiology, phylogeny and in situ evidence for bacterial and archaeal nitrifiers in the marine sponge Aplysina aerophoba". Environmental Microbiology, 10(11): 2942–2955. doi:10.1111/j.1462-2920.2008.01582.x.
- Radax, R., Hoffmann, F., Rapp, H.T., Leininger, S. and Schleper, C. (2012) "Ammonia‐oxidizing archaea as main drivers of nitrification in cold‐water sponges". Environmental Microbiology, 14(4): 909_923. doi:10.1111/j.1462-2920.2011.02661.x.
- Zhang, F., Blasiak, L.C., Karolin, J.O., Powell, R.J., Geddes, C.D. and Hill, R.T. (2015) "Phosphorus sequestration in the form of polyphosphate by microbial symbionts in marine sponges". Proceedings of the National Academy of Sciences, 112(14): 4381–4386. doi:10.1073/pnas.1423768112.
- Colman, A.S. (2015) "Sponge symbionts and the marine P cycle". Proceedings of the National Academy of Sciences, 112(14): 4191–4192. doi:10.1073/pnas.1502763112.
- Simister, R., Taylor, M.W., Tsai, P., Fan, L., Bruxner, T.J., Crowe, M.L. and Webster, N. (2012) "Thermal stress responses in the bacterial biosphere of the Great Barrier Reef sponge, Rhopaloeides odorabile. Environmental Microbiology, 14(12): 3232–3246. doi:10.1111/1462-2920.12010.
- Morrow, K.M., Bourne, D.G., Humphrey, C., Botté, E.S., Laffy, P., Zaneveld, J., Uthicke, S., Fabricius, K.E. and Webster, N.S. (2015) "Natural volcanic CO 2 seeps reveal future trajectories for host–microbial associations in corals and sponges". The ISME Journal, 9(4): 894–908. doi:10.1038/ismej.2014.188.
- Ribes, M., Calvo, E., Movilla, J., Logares, R., Coma, R. and Pelejero, C. (2016) "Restructuring of the sponge microbiome favors tolerance to ocean acidification. Environmental Microbiology Reports, 8(4): 536–544. doi:10.1111/1758-2229.12430.
- Lesser, M.P., Fiore, C., Slattery, M. and Zaneveld, J. (2016) "Climate change stressors destabilize the microbiome of the Caribbean barrel sponge, Xestospongia muta". Journal of Experimental Marine Biology and Ecology, 475: 11–18. doi:10.1016/j.jembe.2015.11.004.
- Chevrette, M.G., Bratburd, J.R., Currie, C.R. and Stubbendieck, R.M. (2019 "Experimental Microbiomes: Models Not to Scale". mSystems, 4(4): e00175-19. doi:10.1128/mSystems.00175-19.
| Groups |  | |
|---|---|---|
| Microbiology | ||
| Ecology | ||
| Plants | ||
| Marine | ||
| Human related | ||
| Techniques | ||
| Other | ||
| ||