Paleolimnology
Paleolimnology (Greek: paleon=old, limne=lake, logos=study) is a scientific subdiscipline closely related to both limnology and paleoecology. Palaeolimnological studies are concerned with reconstructing the paleoenvironments of inland waters (lakes and streams;
| Part of a series on |
| Paleontology |
|---|
 |
|
Paleontology Portal Category |
freshwater, brackish, or saline) – and especially changes associated with such events as climatic change, human impacts (e.g., eutrophication, or acidification), and internal ontogenic processes.
Paleolimnological studies are commonly based on meticulous analyses of sediment cores, including the physical, chemical and mineralogical properties of sediments, and diverse biological records (e.g., fossil diatoms, cladocera, ostracodes, molluscs, pollen, pigments, or chironomids). One of the primary sources for recent paleolimnological research is the Journal of Paleolimnology.
History
Lake ontogeny
Most early paleolimnological studies focused especially on the biological productivity of lakes, and the role of internal lake processes in directing lake development. Although Einar Naumann had speculated that the productivity of lakes should gradually decrease due to leaching of catchment soils, August Thienemann suggested that the reverse process likely occurred. Early midge records seemed to support Thienemann's view.[1]
Hutchinson & Wollack suggested that following an initial oligotrophic stage lakes would achieve and maintain a trophic equilibrium. They also stressed parallels between the early development of lake communities, and the sigmoid growth phase of animal communities – implying that the apparent early developmental processes in lakes were dominated by colonization effects, and lags due to the limited reproductive potential of the colonising organisms.[1]
In a classic paper, Raymond Lindeman[2] outlined a hypothetical developmental sequence, with lakes progressively developing through oligotrophic, mesotrophic, and eutrophic stages, before senescing to a dystrophic stage and filling completely with sediment. A climax forest community would eventually be established on the peaty fill of the former lake basin. These ideas were further elaborated by Ed Deevey,[3] who suggested that lake development was dominated by a process of morphometric eutrophication. As the hypolimnion of lakes gradually filled with sediments, oxygen depletion would promote the release of iron-bound phosphorus to the overlying water. This process of internal fertilization would stimulate biological productivity, further accelerating the in-filling process.[4]
Deevey and Lindemann's ideas were widely, if uncritically, accepted. Although these ideas are still widely held by some limnologists, they were effectively refuted in 1957 by Deevey's student Daniel A. Livingstone.[5] Mel Whiteside[6] also criticized Deevey and Lindemann's proposal, and palaeolimnologists now consider that a host of external factors are equally or more important as regulators of lake development and productivity. Indeed, late-glacial climatic oscillations (e.g., the Younger Dryas) appear to have been accompanied by parallel changes in productivity, illustrating that 1) lake development is not a unidirectional process, and 2) climatic change can have a profound effect on lake communities.
Anthropogenic eutrophication, acidification, and climate change
Interest in paleolimnology eventually shifted from esoteric questions of lake ontogeny to applied investigations of human impact. Torgny Wiederholm and Bill Warwick, for example, used chironomid fossils to assess the impacts of increased nutrient loading (anthropogenic eutrophication) on lake communities. Their studies revealed pronounced changes in the bottom fauna of North American and European lake as a consequence of severe oxygen depletion in the eutrophic lakes.
From 1980 to 1990 the primary focus of paleolimnologists efforts shifted to understanding the role of human impacts (acid rain) versus natural processes (e.g. soil leaching) as drivers of pH change in northern lakes.[7] The pH-sensitivity of diatom communities had been recognized as early as the 1930s, when Friedrich Hustedt developed a classification for diatoms based on their apparent pH preferences. Gunnar Nygaard subsequently developed a series of diatom pH indices. By calibrating these indices to pH, Jouko Meriläinen introduced the first diatom-pH transfer function. Using diatom and chrysophyte fossil records, research groups were able to clearly demonstrate that many northern lakes had rapidly acidified in response to increased industrialization and emissions.[8] Although lakes also showed a tendency to acidify slightly during their early (late-glacial) history, the pH of most lakes had remained stable for several thousand years prior to their recent human-driven acidification.
In recent years palaeolimnologists have recognised that climate is a dominant force in aquatic ecosystem processes, and have begun to use lacustrine records to reconstruct paleoclimates. Sensitive records of climate change have been developed from a variety of indicators including, for example, paleotemperature reconstructions derived from chironomid fossils,[9] and palaeosalinity records inferred from diatoms.[10]
Recent studies in Arctic show that changes in biodiversity are largely due to arctic warming rather than other associated factors such as human alteration and acidification.[11] In the Himalayan region water bodies are not only affected by the anthropogenic disturbances but also impacted by the different types of pollutants that are transferred to this area over a longer distance. Therefore, it is vital to understand all the associated factors acting on the aquatic biodiversity while analyzing the impact of climate change over the past years with the help of lake sediments.[12] It is also important to consider that the impact and intensity of climate change effects varies depending on the ecosystem's sensitivity to change when assessing climate change from a paleolimnological perspective.[13]
Paleoclimate Proxies
.jpg.webp)
Paleoclimatology (or the study of past climates) uses proxy data in order to relate elements collected in modern-day samples to climatic conditions of the past. In paleolimnology, proxy data refers to preserved or fossilized physical markers which serve as substitutes for direct meteorological measurements.[14]
Sediment cores
Sediment cores are one of the primary tools for studying paleolimnology because of the role lake and river sediments play in preserving biological information.[15] Paleolimnologists collect sediment cores and observe various proxy indicators in order to reconstruct past limnology of an area.[15] Proxy data derived from sediment cores includes geochemical markers and isotope data as well as fossilized pollen, charcoal, diatoms, chironomids, and other organic matter.[15] These proxies show diagnostic distributions and characteristics that can indicate past limnological conditions. In order to calibrate the proxy data derived from sediment cores, the new core is compared to a group of around 40 or more calibration lakes.[15] These calibration lakes offer a baseline set of limnological conditions to which the data from the new core is compared.[15]This allows researchers to assess key differences in the limnological conditions of the lake from which the core is derived. Lake sediment cores in particular offer a more comprehensive analysis of an area because of the continual accumulation of sediment as well as other organic matter like pollen and charcoal. Sediment cores can also be dated quite accurately, often allowing for lake histories to be reconstructed in chronological sequence.[16]
Pollen records
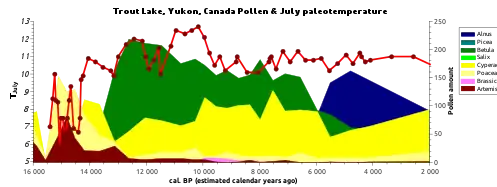
Pollen and spores of terrestrial vegetation around a lake are often found within sediment cores and can be analyzed within a lab setting to determine the family, genus, or species to which the pollen grains belong.[17]Examination of the distribution of these pollen grains and the overall taxonomic assemblage can offer insight into the historical distribution of vegetation around the lake.[18][17] Pollen records derived from paleolimnological assessments also allow researchers to track the distribution and density of different vegetation classes across large periods of time and model the plant succession of the surrounding landscape.[19] Several studies have been able to assess transitions in vegetation profiles by examining the relationship between different types of land cover. For instance, an increase in the presence of fern pollen and herbaceous plant pollen coinciding with a decrease in grassland pollen often indicates a major disturbance or significant land clearance.[19]Another trend that can be observed from historical pollen records is rates of soil erosion around the lake, as arboreal pollen rates often increase with soil erosion due to increased pollen levels in surface soils.[18][19]
Vegetation profiles derived from historical pollen analysis are also seen as a key tool in assessing historical climate trends and disturbances. Pollen analysis offers a historical record of vegetation profiles which are sensitive to abrupt changes in climate conditions. Therefore, historical climate events as well as human-induced climate change can shift vegetation profiles relatively rapidly compared to natural transitions in vegetation profiles. For example, the quantity of poplar pollen increased dramatically at the beginning and end of the Younger Dryas period, serving as a biological marker for the time period.[20]Comparing historical vegetation profiles also allows researchers to compare historical vegetation succession between two specific regions and correlate these differences with the corresponding climates of each region.[21] A recent study conducted at Shudu Lake in the Hengduan Mountains of Yunnan, southwestern China was able to correlate changes in humidity and temperature with the development of needleleaf forests, as well as model recent anthropogenic effects on vegetation distribution in the area.[21]
Diatoms
The taxonomic assemblages of diatoms reflect many aspects of the lake temperature, chemical, and nutrient environment. Diatoms are particularly suited to paleolimnology due to their silica-based frustules, which preserve in sufficient condition and in large enough quantities to be extracted from sediment cores and identified down to the species level.[22]Diatoms have also been examined in conjunction with chrysophycean statospores to estimate nutrient conditions of prehistoric temperate lakes.[22]Such estimations can be derived due to the fact that the relative competitiveness of the two algal groups varies depending on the nutrient conditions of their limnic environment. Diatoms show a high degree of success in water with a high nutrient content as opposed to chyrsophytes who are competitively superior in water with a lower nutrient content.[22]Certain species of diatoms also demonstrate a preference for specific pH conditions allowing researchers to estimate the historical pH conditions of a body of water by analyzing the species of diatoms isolated from a sediment core.[23] This makes them well suited for examinations of the impact of acid rain on a specific body of water as Diatom inference techniques are advanced enough to estimate relatively small numeric ranges of nutrient levels and pH values, as well as fluctuations in these measurements over a certain paleolimnological period. [24]
Organic matter analysis
Examinations of the deposition and makeup of organic matter in the sediments of lake has often been utilized in paleolimnological assessments.[25] A variety of factors are taken in consideration when examining deposited organic matter including quantity, origin, and a variety of isotopes and biomarkers.[25] Diagenesis can have significant impacts on these factors, and thus careful consideration of its impacts are required when drawing conclusions from organic matter records.[25]
Quantity
The quantity of organic matter derived from a sediment core can offer a variety of insights into paleolimnological conditions of a body of water. The quantity of organic matter often serves as an indicator for primary productivity levels as well as terrestrial nutrient input.[26] It also serves a bridge between paleolimnology and geochemistry in demonstrating the relationship between lake geochemistry and organic matter deposition. For instance, a study in eastern China found that larger and deeper highstand lakes in warmer more humid climates tended to show higher levels of organic matter deposition than lowland lakes in cooler arid climates.[26] The same study found that the only factor controlling organic matter deposition in the lowland lakes was primary productivity, whereas organic matter deposition in the highland lakes was controlled by a wider array of factors within the lake ecosystem including terrestrial nutrient input and freshwater inflow.[26]
Origin
By deriving the origin of fossilized organic matter, researchers can make assessments about the vegetation profile in and around the lake, as well as gain a better understanding of microbial density within the lake sediments.[25] A key technique in determining the origin of deposited organic matter is to examined the Carbon to Nitrogen ratio (C:N). Aquatic plants are largely non-vascular, which results in their organic matter having a relatively low C:N ratio relative to that of vascular terrestrial plants.[25] This disparity is usually quite large, and although it is lessened by alterations to the C:N ratio during diagenesis, the original disparity is still evident enough to allow researchers to assess the origin of the organic matter in the lake accurately from C:N ratios.[25] This helps researchers determine algal density and terrestrial organic matter inputs during specified historical periods. Several biomarkers also aid in the determination of organic matter origin. Lipid extraction in particular is a very common practice as it can reveal acids and alcohols characteristic of algal plants, as well as diagnostic lipids generated in the waxy cuticle of terrestrial plants.[25] Lignin phenols also serve as key biomarkers, helping researchers distinguish the source, plant type, tissue type, and age of organic matter.[27] Lignin is particularly useful in distinguishing between angiosperms and gymnosperms as well as between woody and non-woody tissue types, helping researchers further develop their knowledge of the surrounding vegetation.[27] It is also important to note that both biomarkers and the C:N ratio can be altered by microbial interactions, some of which can serve as metrics for measuring microbial density, further adding to the breadth of paleolimnological information that can be derived from examinations of organic matter.[25]
Carbon Isotope Analysis
Three main carbon fixation pathways exist for plants that end up as deposited organic matter: the C3 ,C4, and CAM pathways which all contain slightly different carbon isotope shifts. These shifts further diversify when examining the differences in theses pathways between terrestrial and aquatic plants.[25] However, the impact of microbial degradation and food web interactions diminishes the usefulness of carbon isotopes when differentiating the origin of organic matter.[25]Nonetheless, total quantity of carbon isotopes can reveal characteristics of lake biochemistry, as periods of time characterized by excessive nutrient cycling generally demonstrate lower carbon isotope loads in deposited organic matter.[25] Additionally, heavier carbon isotope shifts are sometimes observed in organic matter deposited during periods of time with drier conditions.[28]
Nitrogen Isotope Analysis
Nitrogen, like carbon shows characteristic isotope shifts depending on its fixation pathway that can be used to assess certain paleolimnological indices. However, also like carbon, a variety of factors play into the nitrogen isotope composition of lake sediments making assessments derived from this method inherently speculative to a degree.[29]In particular, δ15N values can vary based on productivity levels in aquatic ecosystems. A study that reconstructed lake conditions of Lago Taypi in Cordillera Real, Bolivia found that when Nitrogen served as the limiting nutrient in this instance, levels of nitrogen fixing algae significantly rose.[29] These algal groups produce δ15N values that closely aligned with those of atmospheric N2 allowing the researchers to draw conclusions about nutrient cycling and productivity in the lake by examining specific nitrogen isotopes of their sediment cores.[29] Furthermore, in examinations of historic eutrophication trends, δ15N values can be used to differentiate human-driven nitrogen loads from natural inputs, allowing researchers to track the impact of agriculture on historic nitrogen trends.[30]Human and animal waste as well as synthetic fertilizers all have diagnostic isotopic shifts that allow researchers to characterize specific nitrogen inputs and track potential human-induced changes in nutrient flux using δ15N measurements.[30]
Chironomids

Chironomids as a Paleoclimate Proxy
Lake deposits have a rich diversity of fossilized insects that trace back to middle Paleozoic era further increasing in abundance during the Quaternary period. Among the diverse array of aquatic invertebrates, different families of aquatic fly larvae can be extracted from sediments of the Quaternary era. Among them, Chironomids, two-winged flies that belong to the family Chironomidae, are of greatest ecological importance due to their diverse feeding habitat and their role as an important component of the food web. Chironomids complete their larval stage in the water with their adult life stage outside of the water only lasting a very short time. During their larval stages, Chironomids play an important role in the degradation of material in the aquatic ecosystem and are an important part of the food web.[31] Ecologically they are considered bottom dwellers and are very responsive to any fluctuation in the surrounding environment. Their head capsule and feeding structures are quite commonly fossilized in lake sediments,[32] allowing them to serve as a valuable paleoclimate proxy.
Factors influencing Chironomid distribution and abundance
One of the major factors that affects chironomid distribution is the climate conditions at the local, regional, and global scale. Changes in these conditions are preserved over large periods of time as a fossil record. Through paleolimnological methods including chironomid asessment, these changes can be traced out to assess future climate change. Chironomids are very responsive to any fluctuation in the surrounding environment, making them good indicators of a variety of factors including salinity, water depth, stream flow, aquatic productivity, oxygen level, lake acidification, pollution, temperature, and overall ecosystem health. Chironomid distribution can be easily related to those changing factors using a transfer function to connect a particular group of organisms to a specific environmental variable.
A variety of disparate factors have been influencing the abundance and distribution pattern of chironomids in recent history. Therefore, it is important to be careful when making broader interpretations from their fossil records. Several debates in recent history have discussed the impact of temperature on chironomid abundance and diversity along with other associated factors. Accurate interpretations of chironomid fossil records must consider a wide array of associated factors within the ecosystem. In order to understand the different forces that have been affecting the fossil data of a lake, it important to reconstruct physical, chemical and the nutrient content of the lakes that actually shapes the communities. Their distribution and abundance are highly influenced by the combination of human disturbance and change in the climate. Both of them influence the catchment area that resulted into the changing of vegetation, hydrology, and nutrients cycle. Any change at the regional level especially the temperature affects the water quality at the local scale and then at the end it affect the habitat that is species-specific.[32]
Chironomids and reconstruction of quantitative change in Holocene climate
Several factors are considered as controlling factors of chironomid distribution and abundance. Researchers assessing chironomid distribution primarily examine temperature while considering supporting factors such as pH, salinity, nutrient flow and productivity especially in the late Pleistocene/Holocene time period. For many years research has been carried out on the relationship between temperature and chironomids distribution pattern due to the impact of temperature on chironomid emergence. Chironomids are directly and indirectly affected by temperature during the whole life cycle including their larval emergence, growth, feeding and reproduction.[32] According to Eggermont and Heiri the indirect impact of temperature on different physical and chemical aspects determines chironomid distribution and abundance.[9] There is also a strong relationship between the chironomids abundance, emergence and distribution with mean water and air temperatures.[9]According to a research conducted in high altitude lake Lej da la Tscheppain Switzerland, seasonal temperature reconstruction can be done with the help of independent chironomids and diatoms.[33] Any change in the assemblage of chironomids reflects change in the temperature and duration of ice cover of that specific water body due to climate change. According to their findings, chironomids respond mostly to the change in the summer temperature, so seasonal variation in temperature can be inferred from different cores of the sediments.[33]
Use of Chironomids in assessments of anthropogenic Climate Change
According to the fifth IPCC report (2014), a key factor in the shaping of aquatic biodiversity is the progression of human-induced climate change.[34] Macroinvertebrates, especially chironomids have been considered an important indicator of past climate change, especially with regards to temperature. There is a strong correlation between the chironomids assemblage and the water temperature, lake depth, salinity and nutrients concentrations. Lake water levels over the past time period can be inferred from the chironomid assemblage due to this correlation. Therefore, the impact of climate change on lake water levels can be related to changes in the pattern of chironomid distribution and abundance. This strong correlation helps to build the information about the evaporation and precipitation profiles of the lake in the past. Past climatic conditions are reconstructed based on the paleolimnology with the help of different fossilized records especially the lake sediments that helps differentiate regional and local climate change.[31]
References
- Walker, Ian R. (1987-01-01). "Chironomidae (Diptera) in paleoecology". Quaternary Science Reviews. 6 (1): 29–40. doi:10.1016/0277-3791(87)90014-X. ISSN 0277-3791.
- Lindeman, Raymond L. (1991-03-01). "The trophic-dynamic aspect of ecology". Bulletin of Mathematical Biology. 53 (1): 167–191. doi:10.1007/BF02464428. ISSN 1522-9602. S2CID 189882966.
- Deevey, E. S., Jr. 1955. The obliteration of the hypolimmon. Mem. Ist. Ital. Idrobiol., Suppl 8, 9-38.
- Walker, I. R. 2006. Chironomid overview. pp.360–366 in S.A. EIias (ed.) Encyclopedia of Quaternary Science, Vo1. 1, Elsevier, Amsterdam
- Livingstone, D.A. 1957. On the sigmoid growth phase in the history of Linsley Pond. American Journal of Science 255: 364–373.
- Whiteside. M. C. 1983. The mythical concept of eutrophication. Hydrobiologia 103, 107–111.
- Battarbee, R. W. 1984. Diatom analysis and the acidification of lakes. Philosophical Transactions of the Royal Society of London 305: 451–477.
- Battarbee, R. W.; Thrush, B. A.; Clymo, R. S.; Le Cren, E. D.; Goldsmith, P.; Mellanby, K.; Bradshaw, A. D.; Chester, P. F.; Howells, Gwyneth D.; Kerr, A. (1984). "Diatom Analysis and the Acidification of Lakes [and Discussion]". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 305 (1124): 451–477. doi:10.1098/rstb.1984.0070. ISSN 0080-4622. JSTOR 2396098.
- Eggermont, Hilde; Heiri, Oliver (2012-05-01). "The chironomid-temperature relationship: expression in nature and palaeoenvironmental implications". Biological Reviews. 87 (2): 430–456. doi:10.1111/j.1469-185X.2011.00206.x. PMID 22032243. S2CID 29430830.
- Wilson, Susan E.; Cumming, Brian F.; Smol, John P. (1994-12-01). "Diatom-salinity relationships in 111 lakes from the Interior Plateau of British Columbia, Canada: the development of diatom-based models for paleosalinity reconstructions". Journal of Paleolimnology. 12 (3): 197–221. doi:10.1007/BF00678021. ISSN 1573-0417. S2CID 140684939.
- Rahel, Frank J.; Olden, Julian D. (2008-06-01). "Assessing the Effects of Climate Change on Aquatic Invasive Species". Conservation Biology. 22 (3): 521–533. doi:10.1111/j.1523-1739.2008.00950.x. ISSN 0888-8892. PMID 18577081.
- Sharma, C.M; Sharma, S; Gurung, S; Bajracharya, R.M; Jüttner, I; Pradhan, N.S (2009). "Global Climatic Change and High Altitude Lakes: Impacts on Aquatic Biodiversity and Pollution Status". Natural Resources Management: Reviews and Research in the Himalayan Watersheds. 44 (977): 103–122.
- Beniston, Martin (2003-07-01). "Climatic Change in Mountain Regions: A Review of Possible Impacts". Climatic Change. 59 (1): 5–31. doi:10.1023/A:1024458411589. ISSN 1573-1480. S2CID 152554891.
- "What Are "Proxy" Data? | National Centers for Environmental Information (NCEI) formerly known as National Climatic Data Center (NCDC)". www.ncdc.noaa.gov. Retrieved 2020-10-20.
- Tracking environmental change using lake sediments. [Volume 5], Data handling and numerical techniques. Birks, H. J. B. (Harry John Betteley). Dordrecht: Springer. 2012. ISBN 978-94-007-2745-8. OCLC 786034304.CS1 maint: others (link)
- Björck, Svante; Wohlfarth, Barbara (2001), Last, William M.; Smol, John P. (eds.), "14C Chronostratigraphic Techniques in Paleolimnology", Tracking Environmental Change Using Lake Sediments: Basin Analysis, Coring, and Chronological Techniques, Developments in Paleoenvironmental Research, Dordrecht: Springer Netherlands, pp. 205–245, doi:10.1007/0-306-47669-x_10, ISBN 978-0-306-47669-3, retrieved 2020-11-17
- Widyani, K; Ridwansyah, I; Syahrulyati, T (2020-08-11). "Paleolimnology analysis: the reconstruction of Lake Maninjau with pollen as the proxy". IOP Conference Series: Earth and Environmental Science. 535: 012005. doi:10.1088/1755-1315/535/1/012005. ISSN 1755-1315.
- Li, Yue-Cong; Author, Qing-Hai Xu Corresp; Yang, Xiao-Lan; Chen, Hui; Lu, Xin-Miao (2005-09-01). "Pollen‐vegetation relationship and pollen preservation on the Northeastern Qinghai‐Tibetan Plateau". Grana. 44 (3): 160–171. doi:10.1080/00173130500230608. ISSN 0017-3134. S2CID 84621596.
- Msaky, Emma S.; Livingstone, Daniel; Davis, Owen K. (2005-07-01). "Paleolimnological investigations of anthropogenic environmental change in Lake Tanganyika: V. Palynological evidence for deforestation and increased erosion". Journal of Paleolimnology. 34 (1): 73–83. doi:10.1007/s10933-005-2398-0. ISSN 1573-0417. S2CID 129222977.
- Gajewski, Konrad; Viau, Andre E. (2011), "Abrupt Climate Changes During the Holocene Across North America From Pollen and Paleolimnological Records", Abrupt Climate Change: Mechanisms, Patterns, and Impacts, Washington, D. C.: American Geophysical Union, pp. 161–171, doi:10.1029/2010gm001015, ISBN 978-0-87590-484-9, retrieved 2020-10-20
- Yao, Yi-Feng; Song, Xiao-Yan; Wortley, Alexandra H.; Wang, Yu-Fei; Blackmore, Stephen; Li, Cheng-Sen (2017-02-09). "Pollen-based reconstruction of vegetational and climatic change over the past ~30 ka at Shudu Lake in the Hengduan Mountains of Yunnan, southwestern China". PLOS ONE. 12 (2): e0171967. doi:10.1371/journal.pone.0171967. ISSN 1932-6203. PMC 5300144. PMID 28182711.
- Smol, John P. (1985-04-01). "The ratio of diatom frustules to chrysophycean statospores: A useful paleolimnological index". Hydrobiologia. 123 (3): 199–208. doi:10.1007/BF00034378. ISSN 1573-5117. S2CID 44486649.
- Chen, Su-Hwa; Wu, Jiunn-Tzong (1999-09-01). "Paleolimnological environment indicated by the diatom and pollen assemblages in an alpine lake in Taiwan". Journal of Paleolimnology. 22 (2): 149–158. doi:10.1023/A:1008067928365. ISSN 1573-0417. S2CID 67789428.
- Köster, Dörte; Pienitz, Reinhard (2006-02-01). "Seasonal Diatom Variability and Paleolimnological Inferences – A Case Study". Journal of Paleolimnology. 35 (2): 395–416. doi:10.1007/s10933-005-1334-7. ISSN 0921-2728. S2CID 33863663.
- Meyers, Philip A.; Ishiwatari, Ryoshi (1993-09-01). "Lacustrine organic geochemistry—an overview of indicators of organic matter sources and diagenesis in lake sediments". Organic Geochemistry. 20 (7): 867–900. doi:10.1016/0146-6380(93)90100-P. hdl:2027.42/30617. ISSN 0146-6380.
- Ma, Yiquan; Fan, Majie; Lu, Yongchao; Liu, Huimin; Hao, Yunqing; Xie, Zhonghuai; Liu, Zhanhong; Peng, Li; Du, Xuebin; Hu, Haiyan (2016-10-01). "Climate-driven paleolimnological change controls lacustrine mudstone depositional process and organic matter accumulation: Constraints from lithofacies and geochemical studies in the Zhanhua Depression, eastern China". International Journal of Coal Geology. 167: 103–118. doi:10.1016/j.coal.2016.09.014. ISSN 0166-5162.
- Kuliński, Karol; Święta-Musznicka, Joanna; Staniszewski, Andrzej; Pempkowiak, Janusz; Latałowa, Małgorzata (2007-11-01). "Lignin degradation products as palaeoenvironmental proxies in the sediments of small lakes". Journal of Paleolimnology. 38 (4): 555–567. doi:10.1007/s10933-007-9090-5. ISSN 1573-0417. S2CID 129619352.
- Talbot, Michael R.; Johannessen, Truls (1992-05-01). "A high resolution palaeoclimatic record for the last 27,500 years in tropical West Africa from the carbon and nitrogen isotopic composition of lacustrine organic matter". Earth and Planetary Science Letters. 110 (1): 23–37. doi:10.1016/0012-821X(92)90036-U. ISSN 0012-821X.
- Abbott, Mark B.; Wolfe, Brent B.; Aravena, Ramón; Wolfe, Alexander P.; Seltzer, Geoffrey O. (2000-12-01). "Holocene hydrological reconstructions from stable isotopes and paleolimnology, Cordillera Real, Bolivia". Quaternary Science Reviews. 19 (17): 1801–1820. doi:10.1016/S0277-3791(00)00078-0. ISSN 0277-3791.
- Botrel, Morgan; Gregory-Eaves, Irene; Maranger, Roxane (2014-12-01). "Defining drivers of nitrogen stable isotopes (δ15N) of surface sediments in temperate lakes". Journal of Paleolimnology. 52 (4): 419–433. doi:10.1007/s10933-014-9802-6. ISSN 0921-2728. S2CID 130543064.
- Long-term environmental change in Arctic and Antarctic lakes. Pienitz, R. (Reinhard), Douglas, Marianne S. V., Smol, J. P. (John P.). Dordrecht: Springer. 2004. ISBN 978-1-4020-2126-8. OCLC 234238083.CS1 maint: others (link)
- Cohen, Andrew S., 1954- (2003). Paleolimnology : the history and evolution of lake systems. Oxford: New York. ISBN 978-0-19-535089-0. OCLC 59006962.CS1 maint: multiple names: authors list (link)
- von Gunten, Lucien; Heiri, Oliver; Bigler, Christian; van Leeuwen, Jacqueline; Casty, Carlo; Lotter, André F.; Sturm, Michael (2008-04-01). "Seasonal temperatures for the past ∼400 years reconstructed from diatom and chironomid assemblages in a high-altitude lake (Lej da la Tscheppa, Switzerland)". Journal of Paleolimnology. 39 (3): 283–299. doi:10.1007/s10933-007-9103-4. ISSN 1573-0417. S2CID 53007053.
- Climate change 2014 : synthesis report. Pachauri, R. K.,, Mayer, Leo., Intergovernmental Panel on Climate Change. Geneva, Switzerland. 2015. ISBN 978-92-9169-143-2. OCLC 914851124.CS1 maint: others (link)