List of transitional fossils
This is a tentative partial list of transitional fossils (fossil remains of groups that exhibits both "primitive" and derived traits). The fossils are listed in series, showing the transition from one group to another, representing significant steps in the evolution of major features in various lineages. These changes often represent major changes in morphology and anatomy, related to mode of life, like the acquisition of feathered wings for an aerial lifestyle in birds, or limbs in the fish/tetrapod transition onto land.
| Part of a series on |
| Paleontology |
|---|
 |
|
Paleontology Portal Category |
.jpg.webp)
Darwin noted that transitional forms could be considered common ancestors, direct ancestors or collateral ancestors of living or extinct groups, but believed that finding actual common or direct ancestors linking different groups was unlikely.[1][2] Collateral ancestors are relatives like cousins in genealogies in which they are not in your direct line of descent but do share a common ancestor (in this case it is a grandparent). This kind of thinking can be extended to groups of life. For instance, the well-known Archaeopteryx is a transitional form between non-avian dinosaurs and birds, but it is not the most recent common ancestor of all birds nor is it a direct ancestor of any species of bird alive today. Rather, it is considered an extinct close evolutionary "cousin" to the direct ancestors. This may not always be the case, though, as some fossil species are proposed to be directly ancestral to others, like how Australopithecus anamensis is most likely to be ancestral to Australopithecus afarensis.[3]
Nautiloids to ammonoids
| Appearance | Taxa | Relationships | Status | Description | Image |
|---|---|---|---|---|---|
| >500 Ma |
Subclass: |
 | |||
| 390 Ma |
Order: |
|
|||
| 370 Ma |
Subclass: |
|
 |
Cephalopods
| The Cephalopod Evolutionary Series | ||||||
|---|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Location | Image |
| 296 Ma |
Genus |
The earliest described octopod. | ||||
| 164 Ma |
Genus: |
A primitive octopod. | ||||
| 164.7 Ma |
Genus: |
An early member of the Vampyromorphida. |
 | |||
| 94.3 Ma |
Genus: |
A primitive octopod. | ||||
Evolution of insects
| Appearance | Taxa | Relationships | Status | Description | Location | Image |
|---|---|---|---|---|---|---|
|
411 Ma |
Genus: |
The world's oldest known insect. |
||||
|
411 Ma |
Genus: |
Early springtail. |
||||
|
300 Ma |
Genus: |
Ancestral to cockroaches, mantids and termites. |
||||
|
316.5 Ma |
Genus: |
A primitive cockroach. |
||||
|
140 Ma |
Genus: |
The earliest known Lepidopteran. |
||||
|
100 Ma |
Genus: Manipulator (insect) |
Possible common ancestor between Cockroaches and Praying Mantises. |
||||
|
92 Ma |
Genus: |
The oldest known species of bee. |
 | |||
|
80 Ma |
Genus: |
The earliest known species of ant. |
 | |||
| 56–34 Ma |
Genus: |
First leaf insect from the fossil record. |
||||
|
52 Ma |
Genus: |
Transitional fossil myrmecophile (social parasite of ant colonies) of the rove beetle subfamily Pselaphinae. |
Evolution of spiders
| The Spider Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 390 Ma |
Genus: |
Previously thought to be the world's oldest spider. | |||
| 165 Ma |
Genus |
The oldest known haplogyne spider. | |||
Invertebrates to fish
| The Invertebrates → Fish Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 523 Ma |
Genus: |
Lancelet-like in appearance. Oldest known ancestor of modern vertebrates
Vertebrate characters
|
| ||
| 504 Ma |
Class: |
Had fin rays, chevron-shaped muscles and a notochord. | 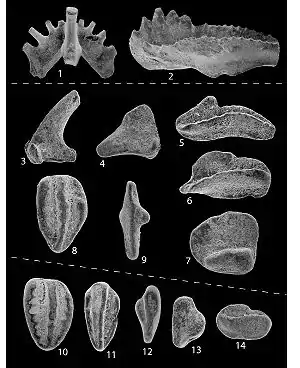 | ||
| 530 Ma |
Genus: |
Appears to have a cranium, thus being a craniate.[4] |  | ||
| 480 to 470 Ma |
Genus: |
Jawless fish | A well armoured jawless fish, resembling a large tadpole in life |  | |
| 422–412 Ma |
Genus: |
An anaspid, ancestral to the jawed vertebrates,[5] | An unarmored, scaly jawless fish |  | |
| 419 Ma |
Genus: |
Oldest known bony fish[6] |  | ||

Chondrichthyes
| The Chondrichthyes Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 370 Ma |
Genus: |
An early primitive shark. |  | ||
| 70–65 Ma |
Genus: |
An early sawfish | |||
| 99–65 Ma |
Genus: |
An early stingray-like skate |  | ||
Bony fish
| The Bony Fish Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 420 Ma |
Genus: |
The earliest-known Actinopterygiian. |
|||
| ??? Ma |
Genus: |
An early relative of the flatfishes, one eye had already migrated towards the body midline. |  | ||
| 48–37 Ma |
Genus: |
The earliest known true flatfish | |||
| 183.7–125.0 Ma |
Genus: |
One of the first teleosts. |  | ||
| 13 Ma |
Genus: |
One of the oldest known seahorses. |  | ||
| 13 Ma |
Genus: |
One of the oldest known seahorses. | 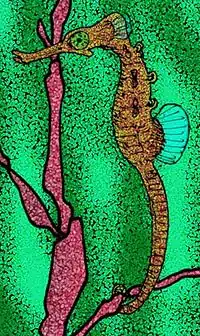 | ||
| 83–70 Ma |
Genus: |
The oldest known lamprid fish |  | ||
| 56–34 Ma |
Genus: |
A primitive sunfish | |||
| 58–55 Ma |
Genus: |
The oldest known member of the catfish family Callichthyidae. |  | ||
| 56–34 Ma |
Genus:
|
A primitive rabbitfish. | |||
| 48–37 Ma |
Genus: |
A primitive perch |  | ||
| 58–55 Ma |
Genus: |
A primitive pomfret |  | ||
| 48–40 Ma |
Genus:
|
An early handfish | |||
| 48–40 Ma |
Genus: |
The oldest known ostraciid boxfish |  | ||
| 48–40 Ma |
Genus: |
The oldest known aracanid boxfish |  | ||
| 48–40 Ma |
Genus:
|
A basal surgeonfish | |||
| 48–40 Ma |
Genus: |
A primitive monodactylid moonyfish |  | ||
| 48–40 Ma |
Genus: |
A primitive monodactylid moonyfish |  | ||
| 48–40 Ma |
Genus: |
A short-snouted ancestor of the modern Moorish Idol. | |||
| 83–65 Ma |
Genus: |
A primitive member of the Tetraodontidae | |||
| 83–65 Ma |
Genus: |
A primitive Perciforme | |||
| 58–55 Ma |
Genus: |
A primitive member of the Zeidae | |||
| 58–55 Ma |
Genus: |
A primitive member of the Zeidae | |||
| ??? Ma |
Genus: |
A primitive member of the Ichthyodectidae |  | ||
| 65 Ma |
Genus: |
A primitive tetraodontid | |||
Fish to tetrapods
| The Fish → Tetrapods Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 416–359 Ma |
Genus: |
An early member of the Tetrapodomorpha, the piscine line leading to tetrapods, Osteolepis is generalised enough to give a fair approximation of the common ancestor of tetrapods and lungfish.[7] | Fish | A small to medium-sized sarcopterygian fish with internal nostrils and pectoral fins stiffened by bony components broadly homologous to the humerus and radius/ulna found in tetrapods.[7] |  |
| 385 Ma |
Genus: |
Belonging to the family Tristichopteridae, a family that form a sister group to Panderichthys and the tetrapods.[7] | Though not on the evolutionary path to tetrapods, Eusthenopteron is of fairly general build and is very well known, serving as an iconic model organism in tetrapod evolution.[8] | A medium-sized, mainly pelagic fish, Eusthenopteron mainly use the pectoral and pelvic fins for navigation, and the tail for propulsion.[8] The fin was of diphycercal, foreshadowing the straightening of the spine and the evolution of a contiguous fin in fish like Panderichthys |  |
| 380 Ma |
Genus: |
Very close to the origin of tetrapods, a "fishapod" elpistostegalian.[7] | Fish | A large, predatory shallow water fish. As common in shallow water fish, the pectoral and pelvic fins were flexible and paddle-like for propulsion.[9] The dorsal and anal fins are lost, the tail fin contiguous.[10] The spiracles were short and wide, indication large amount of oxygen were taken up by the lungs rather than through the gills.[11] |  |
| 375 Ma |
Genus: |
A "fishapod" more tetrapod-like than Panderichthys.[7] | A fish, transitional between fish and the early, fish-like labyrinthodonts.[12][13] | "Fish" with stout, fleshy pectoral fins with a joint between the innermost and the two next bony elements, corresponding to the elbow in higher tetrapods. The cleithrum bone was free of the skull, functioning as anchoring for the pectoral fins, and at the same time allowing for movement of the neck.[13][14] |  |
| 368 Ma |
Genus: |
Analysis of the cranial material shows it was more advanced than Tiktaalik, and together with Obruchevichthys form a sister group to the higher tetrapods.[15] | A fairly fragmentary find, Elginerpeton straddles the fish/tetrapod divide with a mosaic of features resembling Panderichthys, Ichthyostega and Hynerpeton.[15] Probably one of the "fishapods".[16] | Though fragmentary, the find includes a shoulder blade (Cleitrum bone) as well as elements of the limbs, which shows it had comparable limbs Ichthyostega and Hynerpeton, indicating feet rather than fins. |  |
| 365 Ma |
Genus: |
Known only from fragmentary remains, mostly a lower jaw, Ventastega is morphologically midway between Tiktaalik and Acanthostega/Ichthyostega.[17] | Possibly oldest animal to have feet rather than fins.[17] | A large, dorso-ventrally flattened predatory fish with a well armoured labyrinthodont-like skull. While the fins themselves has not been found, the shoulder girdle is essentially similar to that of Acanthostega, indicating it too had feet rather than fins.[17] |  |
| 365 Ma |
Genus: |
Together with Ichthyostega the sole early labyrinthodont known from fairly complete skeletons. It is the oldest animal known to have feet rather than fins, thus making it a true tetrapod and the oldest known unquestionable ichthyostegalian.[18] | First known animal with toes rather than fins. The feet were broad and paddle-like, adapted for movement in water.[19] It retained functional gills in adulthood, behind a fleshy operculum. |  | |
| 365 Ma |
Genus: |
Fairly closely related to Acanthostega. It possibly represent an early (and ultimately unsuccessful) line adapted to moving on land by inchworm-like movements. | Together with Acanthostega the sole early labyrinthodont known from fairly complete skeletons. | Early labyrinthodont with polydactylous, paddle-like feet and reinforced vertebrae and neural spines. It probably spent time on land, yet retained gills and a tail with fin rayes. |  |
| 365 Ma |
Genus: |
An advanced ichthyostegalian, it straddle the divide between the fish-like Devonian forms and the more advanced Carboniferous amphibians. It has been suggested it is an early reptil-like amphibian.[20] | A large animal with paddle-like six-toed feet. It did however not have gills in adulthood, and is thus the oldest labyrinthodont known to depend entirely on breathing with its lungs.[21] |  | |
| 360 Ma |
Genus: |
While known only from fragmentary remains, it is more advanced than Ichthyostega. | Early labyrinthodont amphibian | A large, basically salamander-like creature. The shoulder girdle was powerful, indicating it was a competent walker.[22] |  |
| 359–345 Ma |
Genus: |
Hailing from the fossil-poor Romer's Gap, Pederpes may be ancestral to the higher labyrinthodonts. | Intermediate between the earlier Ichthyostegalian and the later, more advanced labyrinthodonts. | Despite an extra toe on the forelimbs, Pederpes had limbs that terminated in feet adapted primarily for walking rather than paddles for combined swimming and walking like the earlier groups.[23] | |
| 295 Ma |
Genus: |
The Temnospondyli are derived paleozoic amphibians, possibly ancestral to modern amphibians | A "classical" temnospondyl, an advanced labyrinthodont group. | One of the best known labyrinthodonts, Eryops combines the large, flat skull and short limbs typical of the group. | 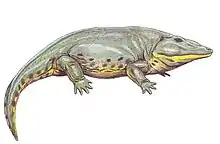 |
| The Labyrinthodontia → Lissamphibia Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 290 Ma |
Genus: |
Colloquially referred to as a "frogamander" due to this taxon being both chronologically and morphologically basal to both anurans and salamanders | One of the first transitional fossils towards modern amphibians (Lissamphibia).[24] | Primitive traits
Derived traits
|
|
| 250 Ma |
Genus: |
Intermediate between generalized amphibians and derived frogs | Early "almost frog" transitional amphibian | Primitive traits
Derived traits
|
|
| 190 Ma |
Genus: |
Another transitional form which could be properly classified as a frog | An intermediate form which may replace Triadobatrachus as the "ultimate" ancestor of anurans | Primitive traits
Derived traits
|
 |
| 213–188 Ma |
Genus: |
A derived fossil frog completing the series of transitional fossils between early amphibians and modern anurans | The oldest "true" frog[25] | Primitive traits
Derived traits
|
|
| 210 Ma |
Genus: |
Intermediate between basal amphibians and caecilians | An early caecilian | Primitive traits
Derived traits
|
|




Amphibians to amniotes
| The Amphibians → Reptiles Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 326–318 Ma |
Genus: |
One of the early reptile-like amphibians | Amphibian | A large, somewhat lizard-like labyrinthodont with a deep skull, laterally placed eyes and five digits to each foot. |  |
| ??? Ma |
Genus: |
The order Diadectomorpha is the sister group of the amniotes. | The Limnoscelis was originally described as a "cotylosaur" (early reptiles) together with the other diadectomorphans. Today the large-bodied diadectomorphs are thought to have had a larval stage, falling close to, but just outside the amphibian/reptile divide. | A large, predatory reptile-like amphibian. The limbs are extremely heavily built, indicating it fed on slow moving prey. | -RedpathMuseumMontreal-June6-08.png.webp) |
| ??? Ma |
Genus: |
Uncertain phylogeny, possibly a Seymouriamorph or Diadectomorph[26][27] | Amphibian | A medium-sized, probably herbivorous animal |  |
| 350 Ma |
Genus: |
Uncertain phylogenetic position. Westlothiana may be a small-bodied diadectopmorph, falling just outside the amphibian/reptile divide | Originally described as the first reptile, it is now considered an advanced reptile-like amphibian. | Small, probably insectovorious animal. The body and tail was long, the limbs small, somewhat like a modern skink. | |
| 320–305 Ma |
Genus: |
Possibly allied to the Diadectomorpha, or belonging to a sister group to Diadectomorpha and Amniota[28] | Likely an amphibian[28] | Smallish, likely carnivorous.[29] |  |
| 340 Ma |
Genus: |
The fragmentary nature of the fossil (it lacks a cranium) makes an exact phylogenetic position hard to establish. | Possibly the first animal with an amniote egg, and thus the first amniote and thus the latest common ancestor to both Synapsids and sauropsids. | Small lizard-like animal, the first known tetrapod to possess claws, indicating it has amniote type skin with scutes.[30] |  |
| 315 Ma |
Genus: |
One of several small, basal reptile genera | Reptile | once thought to be the common ancestor of both synapsids and sauropsids, Hylonomus is now considered a eureptilan creature nested inside sauropsida. |  |
| 312–304 Ma |
Genus: |
One of several small, basal reptile genera | Reptile (most likely a sauropsid) | An early anapsid reptile. In phylogenetic analysis it falls on the sauropsid side, it is thus likely a progenitor of the diapsids |  |
Turtles
| The Turtle Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 240 Ma |
Genus: |
Reptile closely related to turtles. |
| ||
| 220 Ma |
Genus: |
The oldest known turtle. It had a plastron (bottom half of the shell) covering its abdomen. The species broadened ribs are also a key quality as a transitional turtle. It also had teeth and a long tail unlike modern turtles, which earlier ancestors likely had. |
| ||
| 210 Ma |
Genus: |
This species has the oldest known shell consisting fully of a carapace and a plastron. |
 | ||
| 164 Ma |
Genus: |
An evolutionary bridge between early land turtles and sea turtles. |
|||
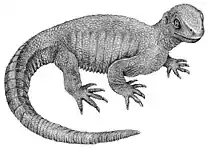

From lizards to snakes
| The Lizard → Snake Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 120 Ma | Genus: | A Basal snake with 4 limbs. | |||
| 95 Ma | Genus: | A basal snake with two hind-limbs containing a hip, knee, and ankle joint. | |||
| 92 Ma |
Genus: |
A transitional form between Cretaceous lizards and limbless snakes retaining distinct, if non-functional, legs.[31] |  | ||
| 90 Ma |
Genus: |
Najash is a key transitional form for snakes. It had a skull with containing a mosaic of features from earlier lizards like having bonier, firmer portions, and a large mouth, mobile joints, and sharp teeth like derived snakes. Najash also had two small, but fully formed back legs. | |||
Lizards
| The Lizard Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 61–58 Ma |
Genus: |
The earliest known chameleon. | |||
| 92 Ma |
Genus: |
A basal mosasauroid from the Upper Cretaceous of North America. | |||
| 71–82 Ma |
Genus: |
One of the earliest Varanoidea. | |||
| 99.42 Ma |
Genus: |
The oldest known gecko | |||
Pterosaurs
| Rhamphorhynchoidea → Pterodactyloidea Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 160 Ma |
Genus: |
Basal to both rhamphorhynchoids and pterodactyloids |  | ||
Archosaurs to dinosaurs
| Appearance | Taxa | Relationships | Status | Description | Image |
|---|---|---|---|---|---|
| 259-252 Ma |
Genus: |
The oldest known archosaur, Archosaurus was one of the largest land reptiles during the Late Permian, about the size of to today's Komodo dragons. It looked somewhat crocodile-like, with sprawling legs, long jaws, powerful neck muscles and a long tail. A distinct proterosuchid trait is the peculiar hook-shaped mouth. |
| ||
| ??? Ma |
Genus: |
 | |||
| ??? Ma |
Genus: |
The oldest known animal on the dinosaur/pterosaur side of the archosaurian tree (the Ornithodira), dating to about 245 million years ago.[32] | A small, lightly built animal. It had a fairly long neck (contrary to the short necked relatives of crocodiles), but ran on all four legs. |  | |
| ??? Ma |
Genus: |
Known from a somewhat fragmentary find, Spondylosoma was possibly an early dinosaur, or near dinosaur.[33] It has however also been classified as a rauisuchian.[34] | |||
| 228 Ma |
Genus: |
A very early representative of the sauropod stem line or perhaps even the Saurischia as a whole.[35][36][37] | A small (1 meter, ~ 10 kg) bipedal carnivore with numerous sharp teeth. It was a swift digigrade runner. The forelimbs were half the length of the hindlimbs and the hands had five fingers |  |
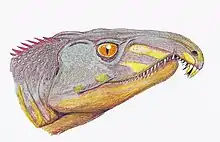
Dinosauria
| Appearance | Taxa | Relationships | Status | Description | Image |
|---|---|---|---|---|---|
| 228 to 216.5 Ma |
Genus: |
The oldest known ornithischian. |
 | ||
| 216–200 Ma |
Genus: |
The most primitive well-known representative of the sauropodomorph dinosaurs. |  | ||
| 160 Ma |
Genus: |
The oldest and most primitive known stegosaur. |  | ||
| 90 Ma |
Genus: |
A basal pachycephalosaur from the late Cretaceous. | |||
| 160 Ma |
Genus: |
A genus of basal ceratopsian dinosaur from the Late Jurassic Period of central Asia. |  | ||
| 160 Ma |
Genus: |
A genus of proceratosaurid tyrannosauroid dinosaur, one of the earliest known examples of the lineage. |  | ||
| 126 Ma |
Genus: |
An early genus of therizinosaur |  | ||
| 208–194 Ma |
Genus: |
One of the most primitive thyreophorans. |  | ||
| 95 Ma |
Genus: |
A possible ancestor of the duck-billed dinosaurs. | |||
| 120 Ma |
Genus: |
A primitive (basal) ornithomimosaur. |
Dinosaurs to birds
| The Dinosaurs → Birds Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 152–151 Ma |
Genus: |
Primitive traits
Derived traits
|
 | ||
| 168–152 Ma |
Genus: |
The find is represented only by a hind leg, but one that is very bird-like. It belonged to a small maniraptoran dinosaur with long, pennaceous feathers on its hind legs and (in all likelihood) arms. | |||
| 161–151 Ma |
Genus: |
Basal troodontid | Although once classified as a bird, Anchiornis is now considered a basal troodontid which bears pennaceous, symmetrical feathers on all four limbs. | Primitive traits
Derived traits
|
 |
| 150–145 Ma |
Genus: |
Known for its mosaic of avian and theropod characteristics Archaeopteryx is both the first primitive bird in the fossil record and one of the first transitional fossils discovered. | Traditionally seen as the first proper bird, though it is not directly ancestral to modern birds.[38] An excellent intermediate form between dinosaurs and birds. Capable of gliding, but lacking alula and keel, it could likely not sustain powered flight. | Primitive traits
Derived traits
|
|
| 120 Ma |
Genus: |
Found in the famous Liaoning province Confuciusornis is the first primitive bird with a pygostyle. | With its short tail and toothless beak, Confuciusornis is very modern looking compared to Archaeopteryx. The toothless beak is however a case of convergent evolution, as more advanced birds retained teeth, illustration the sometimes confusing mosaic evolution of the dinosaur-bird transition. | Primitive traits
Derived traits |
.jpg.webp) |
| 115 Ma |
Genus: |
Primitive bird and possibly a descendant of "urvogels" like Archaeopteryx. First bird to possess an alula. | Plesiomophic traits
Derived traits
|
||
| 93.5–75 Ma |
Genus: |
Considered a close relative to the ancestor to modern birds | A flying bird found in several epochs in the late Cretaceous which still bore teeth, but in most respects very similar to Neornithes. | Primitive traits
Derived traits
|
 |
Bird evolution
| The Bird Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 60–58 Ma |
Genus: |
The earliest-known penguin. |
 | ||
| ??? Ma |
Genus:
|
An early flamingo. | |||
| ??? Ma |
Genus: |
An early gaviiform. | |||
| 55–48 Ma |
Genus: |
An early psittacine. | |||
| ??? Ma |
Genus: |
A basal falconiform. |  | ||
| 50 Ma |
Genus: |
An early apodiform. | |||
Non-mammalian synapsids to mammals
| The Synapsids → Mammals Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 318-315 Ma |
Genus: |
Known from very fragmentary finds, Protoclepsydrops may be the earliest synapsid | A low-slung, lizard-like animal of moderate size. | ||
| 306 Ma |
Genus: |
The oldest undisputed synapsid | Primitive traits
Derived traits
|
 | |
| 297 Ma | Genus: | A primitive member of the Sphenacodontidae, or possibly just outside the group.[40][41] | A pelycosaur-grade synapsid | Derived traits
|
 |
| 265 Ma |
Genus: |
An advanced member of the family Sphenacodontidae, from which the therapsids (advanced synapsids) evolved | A pelycosaur-grade synapsid. At up to 4 meters, Dimetrodon was one of the largest animals of its time. The distinct sail of the back makes it the most recognized synapsid known |
Primitive traits
Derived traits |
 |
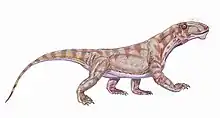
| 267 Ma |
Genus: |
A primitive therapsid. About the size of a large dog, Biarmosuchus was a lightly built and likely fairly agile animal for its size.[45] |
Primitive traits
Derived traits |
 | |
| 247–237 Ma | Genus: | An advanced synapsid | All species of Cynognathus were rather heavyset carnivores about a meter in length and with a sprawling gait and heavy jaws. | Primitive traits
Derived traits |
 |
| 248–245 Ma |
Genus: |
A small bodied relative of the larger Cynognathus. | An advanced non-mammalian cynodont. A burrower that ranged from the size of a marten to a badger. | Primitive traits
Derived traits
|
 |
| 205 Ma |
Genus: |
A smaller, more shrew-like relative of Thrinaxodon and Sinoconodon | An early mammal, possibly representing the earliest lactating animals, but outside the crown group (a mammaliform) | primitive traits
Advanced traits
|
 |
| 125 Ma |
Genus: |
One of the Triconodonts | An early crown group mammal. | Primitive traits
Advanced traits
|
 |
Evolution of mammals
| The Mammal Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 100–104 Ma |
Genus: |
The earliest known monotreme. | |||
|
Genus: |
The oldest metatherian known. |
 | |||
| ?? Ma |
Genus: |
The earliest-known marsupial. |
 | ||
| 164–165 Ma |
Genus: |
 | |||
| 63-50 Ma |
Genus: |
The earliest known proboscidean. |
|||
| 60–55 Ma |
Genus: |
The possible ancestor of the modern order Carnivora. |
 | ||
| 15.97–11.61 Ma |
Genus: |
The earliest known cervid. |
 | ||
| 20–18 Ma |
Genus: |
The earliest known bovid. |
 | ||
| 45–40 Ma |
Genus: |
The oldest camel known, it was also the smallest. |
|||
| ??? Ma |
Genus: |
Suspected to be the ancestor of modern tapirs and rhinoceroses. |
 | ||
| 55.4–48.6 Ma |
Genus: |
Suspected to be the ancestor of modern tapirs. |
 | ||
| 38–33.9 Ma |
Genus: |
The earliest known canid. |
 | ||
| ??? Ma |
Genus:
|
The earliest known lagomorph. |
|||
| 52.5 Ma |
Genus: |
One of the two oldest known monospecific genera of bat. |
|||
| 2 Ma |
Species: |
The earliest known member of the giant panda clade. |
|||
| 63–61.7Ma |
Genus: |
Believed to be the earliest example of a primate or a proto-primate, a primatomorph precursor to the Plesiadapiformes. |  | ||
| 12.5–8.5 Ma |
Genus: |
This genus may have been the ancestor to the modern orangutans. |
 | ||
| 16–8 Ma |
Genus: |
A possible ancestor of living hippopotamids. | |||
| 47 Ma |
Genus: |
The earliest known true (and scaled) pangolin. |  | ||
Early artiodactylans to whales
| The whale Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 55.8 ± 0.2 – 33.9 ± 0.1 Ma |
Genus: |
 | |||
| 50 Ma |
Genus: |
 | |||
| 46 Ma |
Genus: |
 | |||
| 47 Ma |
Genus: |
||||
| 41–33 Ma |
Genus: |
 | |||
| 25 Ma |
Genus: |
 | |||
| 40–34 Ma |
Genus: |
 | |||
| 8–15 Ma |
Genus: |
 | |||
| 26 Ma |
Genus: |
||||
Evolution of sirenians
| The Sirenia Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 50 Ma |
Genus: |
This animal had fully formed hips, and legs bones, meaning it very likely lived a semi-aquatic life similar that of a hippopotamus. |
 | ||
| 40 Ma |
Genus: |
| |||
| ??? Ma |
Genus: |
||||
| 48.6–33.9 Ma |
Genus: |
An evolutionary bridge between primitive land-dwelling sirenians to aquatic sirenians |
 | ||
| ??? Ma |
Genus: |
| |||


Evolution of pinnipeds
| The Pinniped Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 21 to 24 Ma |
Genus: |
Likely had webbed feet, along with some skull similarities to modern pinnipeds. |
| ||
| 23-11 Ma |
Genus: |
A very basal pinniped. |  | ||
| 24–22 Ma |
Genus: |
An early seal, but with more primitive skull and feet. |
| ||
.jpg.webp)

Evolution of the horse
| The Hyracotherium → Equus Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 60–45 Ma |
Genus: |
 | |||
| 40–30 Ma |
Genus: |
 | |||
| 20 Ma |
Genus: |
 | |||
| 17–11 Ma |
Genus: |
 | |||
| 12 Ma |
Genus: |
 | |||
| 1.8–0 Ma |
Genus: |
 | |||
Human evolution
List of human evolution fossils
| The Human Evolutionary Series | |||||
|---|---|---|---|---|---|
| Appearance | Taxa | Relationships | Status | Description | Image |
| 36–32 Ma |
Genus |
The oldest primitive monkey known in the fossil record, dating back before the split between Old and New World monkeys. | Basal to both Old and New World monkeys. | Primitive traits
Derived traits
|
|
| 33 Ma |
Genus |
A Miocene monkey which bridges the gap between the Eocene ancestors of Old World monkeys and Miocene ancestor of Hominoidea. | Tentatively positioned transitional form prior to the Old World monkey/ape split. | Primitive traits
Derived traits
|
 |
| 27–14 Ma |
Genus |
This primate exhibits very ape-like features like its teeth, but much of its post-cranial remains are more similar to monkeys. | Universally accepted to be intermediate between 'ape-like monkeys' such as Aegyptopithecus and later apes including hominids. | Primitive traits
Derived traits
|
 |
| 13 Ma |
Genus: |
A European ape which is considered to be the predecessor of the great apes. | Some objections have been raised to this fossils status due to its location in Spain, but Pierolapithecus is likely a transitional taxon between generalized apes and the lineage which led to great apes. | Pleisomorphic traits
Derived traits
|
|
| 4.4 Ma |
Genus: |
A woodland hominid adapted to quadruped arboreal locomotion, but also for bipedalism. | Intermediate between the last common ancestor of chimps and humans, and the australopithecines. | Primitive traits
Derived traits
|
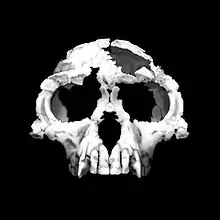 |
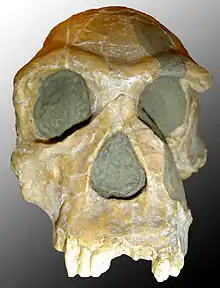
| 4.4–2.0 Ma |
Genus: |
First known genus of fully bipedal apes which are probably ancestral to robust australopiths and the genus Homo. | Intermediate between extinct quadrupedal and bipedal apes. While the relationship between some species are being revised, Australopithecus afarensis is considered to be, by most experts, the ancestor to all later hominids. | Primitive traits
Derived traits
|
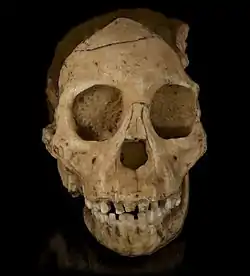 |
| 2.3–1.4 Ma |
Species: |
An early human which is the morphological link between australopithecines and later human species. | Perfect intermediate between early hominids and later humans, possibly ancestral to modern humans. | Primitive traits
Derived traits
|
 |
| 2.0–1.0 Ma |
Species: |
Very successful hominid, which was probably ancestral to both modern humans and neanderthals. Probably the first hominid to leave and successfully colonize territories outside of Africa. | Ancestral to modern humans and neanderthals. | Primitive traits
Derived traits
|
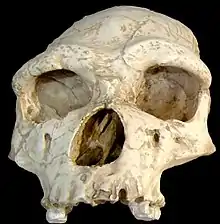 |
| 500 Ka–recent |
Species |
Homo rhodesiensis was the immediate ancestor of modern humans which evidently displaced the neanderthals in Europe and the island 'hobbits' of southeast Asia. H. rhodesiensis evolved from Homo erectus about half a million years ago but still retains some primitive characteristics such as relatively thick bones and molars larger than modern humans. | Ancestral to modern humans. | Primitive traits
Derived traits
|
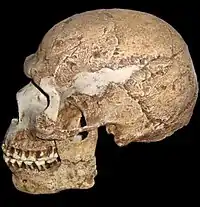 |
See also
References
- Stauffer, RC (1975) Charles Darwin's Natural Selection; being the second part of his big species book written from 1856 to 1858. Cambridge: Cambridge University Press. p. 236.
- Darwin, C. R. 1859. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray. p. 187.
- Delezene, LK; Kimbel, WH (2011). "Evolution of the mandibular third premolar crown in early Australopithecus". Journal of Human Evolution. 60 (6): 711–730. doi:10.1016/j.jhevol.2011.01.006. PMID 21481921.
- Shu, D. G.; Morris, S. C.; Han, J.; Zhang, Z. F.; Yasui, K.; Janvier, P.; Chen, L.; Zhang, X. L.; Liu, J. N.; Li, Y.; Liu, H. -Q. (2003), "Head and backbone of the Early Cambrian vertebrate Haikouichthys", Nature, 421 (6922): 526–529, Bibcode:2003Natur.421..526S, doi:10.1038/nature01264, PMID 12556891, S2CID 4401274
- Ahlberg, Per Erik (2001). Major Events in Early Vertebrate Evolution: Palaeontology, Phylogeny, Genetics, and Development. Washington, DC: Taylor & Francis. p. 188. ISBN 978-0-415-23370-5.
- Zhu, M.; Zhao, W.; Jia, L.; Lu, J.; Qiao, T.; Qu, Q. (2009). "The oldest articulated osteichthyan reveals mosaic gnathostome characters". Nature. 458 (7237): 469–474. Bibcode:2009Natur.458..469Z. doi:10.1038/nature07855. PMID 19325627. S2CID 669711.
- Ahlberg, P. E.; Johanson, Z. (1998). "Osteolepiforms and the ancestry of tetrapods" (PDF). Nature. 395 (6704): 792–794. Bibcode:1998Natur.395..792A. doi:10.1038/27421. S2CID 4430783. Archived from the original (PDF) on 24 November 2014. Retrieved 13 March 2011.
- R. Cloutier (1996). "Taxonomic review of Eusthenopteron foordi.". Devonian Fishes and Plants of Miguasha, Quebec, Canada. Dr. Friedrich Pfeil, München. pp. 487–502.
- Nature: The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion
- Carroll, R. (1995). "Between fish and amphibians". Nature. 373 (6513): 389–390. Bibcode:1995Natur.373..389C. doi:10.1038/373389a0. S2CID 5412926.
- Brazeau, M.D.; Ahlberg, P.E. (2006). "Tetrapod-like middle ear architecture in a Devonian fish". Nature. 439 (7074): 318–321. Bibcode:2006Natur.439..318B. doi:10.1038/nature04196. PMID 16421569. S2CID 4301561.
- John Noble Wilford, The New York Times, Scientists Call Fish Fossil the Missing Link, 5 April 2006.
- Shubin, Neil (2008). Your Inner Fish. Pantheon. ISBN 978-0-375-42447-2.
- "Meet Your ancestor, the Fish that crawled". New Scientist Magazine. Retrieved 7 February 2007.
- Ahlberg, Per E. (1995). "Elginerpeton pancheni and the earliest tetrapod clade". Nature. 373 (6513): 420–425. Bibcode:1995Natur.373..420A. doi:10.1038/373420a0. S2CID 4344655.
- Elginerpeton pacheni at Devonian Times Archived 4 February 2005 at the Wayback Machine
- Ahlberg, Per. E.; Jennifer A. Clack; Ervins Luksevics; Henning Blom; Ivars Zupins (26 June 2008). "Ventastega curonica and the origin of tetrapod morphology". Nature. 453 (7199): 1199–1204. Bibcode:2008Natur.453.1199A. doi:10.1038/nature06991. PMID 18580942. S2CID 4344417. article
- Clack, J. (21 November 2005). "Getting a leg up on land". Scientific American. 293 (6): 100–7. Bibcode:2005SciAm.293f.100C. doi:10.1038/scientificamerican1205-100. PMID 16323697. Archived from the original on 4 November 2006.
- "Acanthostega gunneri," Devonian Times. Archived 24 December 2004 at the Wayback Machine
- Lebedev, O.A. (1984). "The first find of a Devonian tetrapod vertebrate in the USSR". Doklady Akademii Nauk SSSR. Palaeontology (in Russian). 278: 1470–1473.
- Gordon, M.S.; Long, J.A. (2004). "The Greatest Step in Vertebrate History: A Paleobiological Review of the Fish-Tetrapod Transition" (PDF). Physiological and Biochemical Zoology. 77 (5): 700–719. doi:10.1086/425183. PMID 15547790. S2CID 1260442.
- Shubin, Neil (2009). Your Inner Fish: A Journey into the 3.5-Billion-Year History of the Human Body. New York: Vintage. p. 13. ISBN 978-0-307-27745-9.
- Clack, J. A. (2002). "An early tetrapod from 'Romer's Gap'". Nature. 418 (6893): 72–76. Bibcode:2002Natur.418...72C. doi:10.1038/nature00824. PMID 12097908. S2CID 741732.
- Anderson, J. S.; Reisz, R. R.; Scott, D.; Fröbisch, N. B.; Sumida, S. S. (2008). "A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders". Nature. 453 (7194): 515–518. Bibcode:2008Natur.453..515A. doi:10.1038/nature06865. PMID 18497824. S2CID 205212809.
- Estes, R., and O. A. Reig. (1973): "The early fossil record of frogs: a review of the evidence." Pp. 11–63 In J. L. Vial (Ed.), Evolutionary Biology of the Anurans: Contemporary Research on Major Problems. University of Missouri Press, Columbia.
- Moss J.L. (1972). "The Morphology and phylogenetic relationship of the Lower Permian tetrapod Tseajaia campi Vaughn (Amphibia: Seymouriamorpha)". University of California Publications in Geological Sciences. 98: 1–72.
- Berman, D.S.; Sumida, S.S.; Lombard, R.E. (1992). "Reinterpretation of the temporal and occipital regions in Diadectes and the relationship of diadectomorphs". Journal of Paleontology. 66 (3): 481–499. doi:10.1017/S0022336000034028. S2CID 73547163.
- Gauthier J., Kluge, A.G., & Rowe, T. (1988) "The early evolution of the Amniota." In: M. J. Benton (ed.) The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds (1): pp 103–155. Oxford: Clarendon Press.
- Solenodonsaurus on the TOL-web
- R. L. Paton, T. R. Smithson and J. A. Clack, "An amniote-like skeleton from the Early Carboniferous of Scotland", (abstract), Nature 398, 508–513 (8 April 1999)
- "Fossilized Snake With Two Legs Found - Science - redOrbit". 10 April 2008. Retrieved 16 April 2008.
- Nesbitt, S.J.; Sidor, C.A.; Irmis, R.B.; Angielczyk, K.D.; Smith, R.M.H.; Tsuji, L.M.A. (2010). "Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira". Nature. 464 (7285): 95–98. Bibcode:2010Natur.464...95N. doi:10.1038/nature08718. PMID 20203608. S2CID 4344048.
- Langer, M.C. (2004). Basal Saurischia. In: Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.). The Dinosauria (second edition). University of California Press:Berkeley, 25-46. ISBN 0-520-24209-2
- Galton, P.M. (2000). "Are Spondylosoma and Staurikosaurus (Santa Maria Formation, Middle-Upper Triassic, Brasil) the oldest saurischian dinosaurs?". Paläontologische Zeitschrift. 74 (3): 393–423. doi:10.1007/bf02988109. S2CID 140188942.
- R.N. Martinez et al. A basal dinosaur from the dawn of the dinosaur era in southwestern Pangaea. Science, Vol. 331, 14 January 2011, p. 206.
- Kaplan M, "Move over Eoraptor", http://www.nature.com/news, 13-1-2011.
- Apaldetti, C; Martinez, RN; Alcober, OA; Pol, D (2011). "A New Basal Sauropodomorph (Dinosauria: Saurischia) from Quebrada del Barro Formation (Marayes-El Carrizal Basin), Northwestern Argentina". PLOS ONE. 6 (11): e26964. doi:10.1371/journal.pone.0107672. PMC 4178034. PMID 25259845.
- Padian, K. & Chiappe, L.M. (1997): Bird Origins. In: Encyclopedia of Dinosaurs (red. Currie, P.J & Padian, K., Academic Press, San Diego, pp 41–96, ISBN 978-0-12-226810-6
- Chinsamy A.; Martin L.D.; Dobson P. (1998). "Bone microstructure of the diving Hesperornis and the volant Ichthyornis from the Niobrara Chalk of western Kansas". Cretaceous Research. 19 (2): 225–235. doi:10.1006/cres.1997.0102.
- Jörg Fröbisch; Rainer R. Schoch; Johannes Müller; Thomas Schindler; Dieter Schweiss (2011). "A new basal sphenacodontid synapsid from the Late Carboniferous of the Saar-Nahe Basin, Germany" (PDF). Acta Palaeontologica Polonica. 56 (1): 113–120. doi:10.4202/app.2010.0039. S2CID 45410472.
- Michel Laurin (1994). "Re-evaluation of Cutleria wilmarthi, an Early Permian synapsid from Colorado". Journal of Vertebrate Paleontology. 14 (1): 134–138. doi:10.1080/02724634.1994.10011544.
- Romer, A.S.; Price, L.L. (1940). "Review of the Pelycosauria". Special Papers of the Geological Society of America. Geological Society of America Special Papers. 28: 1–538. doi:10.1130/spe28-p1.
- GA Floridesa, Kalogiroua SA; SA; Wrobelb, L Tassoub (2001). "Natural environment and thermal behavior of Dimetrodon limbatus". Journal of Thermal Biology. 26 (1): 15–20. doi:10.1016/S0306-4565(00)00019-X. PMID 11070340.
- Angielczyk, Kenneth D. (June 2009). "Dimetrodon Is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution". Evolution: Education and Outreach. 2 (2): 257–271. doi:10.1007/s12052-009-0117-4.
- White, T. & Kazlev, M. A. (2009): Therapsida: Biarmosuchia: Biarmosuchidae / Eotitanosuchidae, from Palaeos website.
- Ruben, J.A.; Hillenius, W.J.; Kemp, T.S.; Quick, D.E. (2011). "The Evolution of Mammalian Endothermy". In Chinsamy-Turan, A. (ed.). Forerunners of Mammals. Bloomington: Indiana University Press. pp. 272–286. ISBN 978-0-253-35697-0.
- Maier, W.; Heever, J.; Durand, F. (27 April 2009). "New therapsid specimens and the origin of the secondary hard and soft palate of mammals". Journal of Zoological Systematics and Evolutionary Research. 34 (1): 9–19. doi:10.1111/j.1439-0469.1996.tb00805.x.
- Czaplewski, Terry A. Vaughan, James M. Ryan, Nicholas J. (2000). Mammalogy (4th ed.). Fort Worth: Brooks/Cole Thomson Learning. p. 51. ISBN 978-0030250347.
- Ruben, J. A. (1 August 2000). "Selective Factors Associated with the Origin of Fur and Feathers". Integrative and Comparative Biology. 40 (4): 585–596. doi:10.1093/icb/40.4.585.
- "Press release: Paleontologists Discover New Mammal from Mesozoic Era". National Science Foundation. 14 March 2007.
- Zhe-Xi Luo; Chong-Xi Yuan; Qing-Jin Meng; Qiang Ji (25 August 2011). "A Jurassic eutherian mammal and divergence of marsupials and placentals". Nature. 476 (7361): 442–445. Bibcode:2011Natur.476..442L. doi:10.1038/nature10291. PMID 21866158. S2CID 205225806. Electronic supplementary material
External links
- Vuletic.com, Section V: Paleontology – Transitional fossils between every animal group
- Palaeos.com, Palaeos vertebrates starting with lobe-finned fish (very comprehensive)
- Talk.origins.org, FAQ: Transitional vertebrate fossils
- (A few) transitional fossils
