YWHAZ
14-3-3 protein zeta/delta (14-3-3ζ) is a protein that in humans is encoded by the YWHAZ gene on chromosome 8.[5][6] The protein encoded by this gene is a member of the 14-3-3 protein family and a central hub protein for many signal transduction pathways.[6][7] 14-3-3ζ is a major regulator of apoptotic pathways critical to cell survival and plays a key role in a number of cancers and neurodegenerative diseases.[7][8][9][10][11]
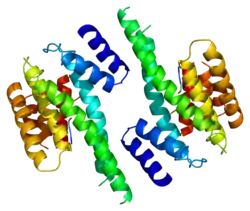



Structure
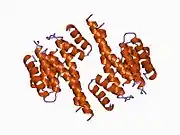
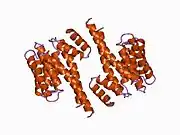
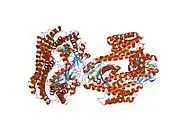
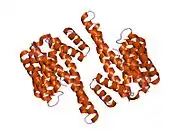
14-3-3 proteins generally form ~30 kDa-long homo- or heterodimers.[12][13] Each of the monomers are composed of 9 antiparallel alpha helices. Four alpha-helices (αC, αE, αG, and αI) form an amphipathic groove that serves as the ligand binding site, which can recognize three types of consensus binding motifs: RXX(pS/pT)XP, RXXX(pS/pT)XP, and (pS/pT)X1-2-COOH (where pS/pT represents phosphorylated serine/threonine). In addition to these primary interactions, the target protein can also bind outside the groove via secondary interactions. In particular, the crystallized structure of 14-3-3ζ forms a cup-shaped dimer when complexed with CBY.[13] The YWHAZ gene encodes two transcript variants which differ in the 5' UTR but produce the same protein.[6]
Function
14-3-3ζ is one of 7 members of the 14-3-3 protein family, which is ubiquitously expressed and highly conserved among plants and mammals.[6][7][11][12] This protein family is known for regulating signal transduction pathways primarily through binding phosphoserine proteins, though it can also bind phosphothreonine proteins and unphosphorylated proteins.[6][7][8][11][14] By extension, 14-3-3 proteins are involved in a wide range of biological processes, including metabolism, transcription, apoptosis, protein transport, and cell cycle regulation.[8][9][11][12][15] This combination of dependence on phosphorylation and widespread biological impact results in dynamic regulation of multiple signalling pathways and allows for cellular adaptation to environmental changes.[8]
In particular, 14-3-3ζ is a key player in regulating cell survival and interacts with many apoptotic proteins, including Raf kinases, BAX, BAD, NOXA, and caspase-2.[8][9] For the most part,14-3-3ζ negatively regulates apoptosis by binding and sequestering BAD and BAX in the cytoplasm, effectively preventing activation of proapoptotic Bcl-2 and Bcl-XL, as well as by preventing NOXA from inhibiting antiapoptotic MCL1.[9] As a result, 14-3-3ζ functions to protect the cell from environmental stresses, such as chemotherapy-induced death, anoikis, growth factor deprivation, and hypoxia. As an example of its dynamic activity, 14-3-3ζ activates autophagy under hypoxic conditions by binding ATG9A, while it prevents autophagy under hyperglycemic conditions by binding Vps34.[8] Furthermore, 14-3-3ζ may regulate glucose receptor trafficking in response to insulin levels through its interaction with IRS1.[6][8]
In addition to cell survival, 14-3-3ζ regulates cell cycle progression through various ligands and processes. For instance, 14-3-3ζ controls cellular senescence by complexing with BIS to chaperone protein folding of STAT3 and activate the signaling pathway.[16] Also, 14-3-3ζ can negatively regulate the G2-M phase checkpoint by binding and sequestering the cyclin-dependent kinases to the cytoplasm, thus inhibiting their activity.[17] Since 14-3-3ζ is predominantly found in the cytoplasm and binds many nuclear proteins, it likely prevents nuclear import by blocking the nuclear localization signal of target proteins.[12] Its localization to both the cytoplasm and nucleus also suggests a role in gene expression, possibly through regulation of transcription factor activity.[9]
Antigenic Function
Emerging literature shows the increased presence of the anti-14-3-3ζ antibodies in several immune dysfunctions, including human vasculitis and cancer.[18][19][20] The antigenic 14-3-3ζ can directly affect T cell differentiation into Th1 and Th17 cells, and thereby promotes IFN-gamma and IL-17 production.[21] The MHC class II presentation of 14-3-3ζ antigen strongly influence IFN-gamma production.[21] The physiological significance of its antigenic role remains unknown
Signaling regulator
Intracellular 14-3-3ζ plays a role in interleukin-17 signaling. IL-17A is a proinflammatory cytokine involved in autoimmune diseases and host defense. The presence of 14-3-3ζ creates a bias in IL-17A signaling outcomes, by promoting the production of IL-6 while suppressing CXCL1.[22]
Clinical Significance
The14-3-3 protein zeta/delta (14-3-3ζ) is a protein (in humans encoded by the YWHAZ gene on chromosome 8) with an important apoptotic constituents. During a normal embryologic processes, or during cell injury (such as ischemia-reperfusion injury during heart attacks and strokes) or during developments and processes in cancer, an apoptotic cell undergoes structural changes including cell shrinkage, plasma membrane blebbing, nuclear condensation, and fragmentation of the DNA and nucleus. This is followed by fragmentation into apoptotic bodies that are quickly removed by phagocytes, thereby preventing an inflammatory response.[23] It is a mode of cell death defined by characteristic morphological, biochemical and molecular changes. It was first described as a "shrinkage necrosis", and then this term was replaced by apoptosis to emphasize its role opposite mitosis in tissue kinetics. In later stages of apoptosis the entire cell becomes fragmented, forming a number of plasma membrane-bounded apoptotic bodies which contain nuclear and or cytoplasmic elements. The ultrastructural appearance of necrosis is quite different, the main features being mitochondrial swelling, plasma membrane breakdown and cellular disintegration. Apoptosis occurs in many physiological and pathological processes. It plays an important role during embryonal development as programmed cell death and accompanies a variety of normal involutional processes in which it serves as a mechanism to remove "unwanted" cells.
As a major hub protein, 14-3-3ζ is involved in various diseases and disorders. For one, 14-3-3ζ plays a central role in cell proliferation and, by extension, tumor progression.[7][10] The protein has been implicated in many cancers, including lung cancer, breast cancer, lymphoma, and head and neck cancer, through pathways such as mTOR, Akt, and glucose receptor trafficking. Notably, it has been associated with chemoresistance and, thus, is a promising therapeutic target for cancer treatment.[8][9][10] So far, it stands to become a prognostic marker for breast cancer, lung cancer, head and neck cancer, and possibly gastric cancer in patients who might require more aggressive treatment.[7] However, no statistically significant relationship was determined in hepatocellular carcinoma.[17]
In addition to cancers, 14-3-3ζ has been implicated in pathogenic infections and neurodegenerative diseases, including Creutzfeldt–Jakob disease, Parkinson’s disease, and Alzheimer’s disease (AD).[11] 14-3-3ζ has been observed to participate in AD through its interaction with tau protein, and its expression is correlated with disease severity.[14]
The human surfactant protein A, an innate immunity molecule (encoded by two genes SFTPA1 and SFTPA2) appears to be binding with the 14-3-3 protein family. Furthermore, inhibition of 14-3-3 was correlated with lower levels of the surfactant protein indicating a relationship between surface and 14-3-3 proteins.[24] Surfactant is an important element in the maintenance of lung and respiratory functions. A lack of surfactant is closely related to respiratory distress syndrome. Preterm neonates who exhibit neonatal respiratory distress syndrome (NRDS) exhibit a deficiency of surfactant. All together, the 14-3-3 protein may have a significant role in respiratory function and NRDS.[25][26]
Interactions
YWHAZ has been shown to interact with:
- IRS1,[6]
- Protein phosphatase 1,[12]
- BIS,[16]
- ATG9A,[8]
- NOXA,[9]
- AKT1,[27]
- BCAR1,[28]
- BAX,[9]
- BAD,[9][29]
- C-Raf,[30][31][32][33][34]
- CDC25B,[35]
- GP1BA,[36][37][38]
- GP1BB,[36][37][39]
- HMGN1,[40]
- IL9R,[41]
- LIMK1,[42]
- P53,[43]
- PRKCE[44]
- PRKCZ,[33][45]
- TNFAIP3,[46][47]
- TSC2,[48]
- Tau protein,[49] and
- VIM.[31]
See also
References
- GRCh38: Ensembl release 89: ENSG00000164924 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000022285 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Tommerup N, Leffers H (April 1996). "Assignment of the human genes encoding 14,3-3 Eta (YWHAH) to 22q12, 14-3-3 zeta (YWHAZ) to 2p25.1-p25.2, and 14-3-3 beta (YWHAB) to 20q13.1 by in situ hybridization". Genomics. 33 (1): 149–50. doi:10.1006/geno.1996.0176. PMID 8617504.
- "Entrez Gene: YWHAZ tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, zeta polypeptide".
- Nishimura Y, Komatsu S, Ichikawa D, Nagata H, Hirajima S, Takeshita H, et al. (April 2013). "Overexpression of YWHAZ relates to tumor cell proliferation and malignant outcome of gastric carcinoma". British Journal of Cancer. 108 (6): 1324–31. doi:10.1038/bjc.2013.65. PMC 3619260. PMID 23422756.
- Weerasekara VK, Panek DJ, Broadbent DG, Mortenson JB, Mathis AD, Logan GN, et al. (December 2014). "Metabolic-stress-induced rearrangement of the 14-3-3ζ interactome promotes autophagy via a ULK1- and AMPK-regulated 14-3-3ζ interaction with phosphorylated Atg9". Molecular and Cellular Biology. 34 (24): 4379–88. doi:10.1128/MCB.00740-14. PMC 4248729. PMID 25266655.
- Liang R, Chen XQ, Bai QX, Wang Z, Zhang T, Yang L, et al. (2014). "Increased 14-3-3ζ expression in the multidrug-resistant leukemia cell line HL-60/VCR as compared to the parental line mediates cell growth and apoptosis in part through modification of gene expression". Acta Haematologica. 132 (2): 177–86. doi:10.1159/000357377. PMID 24603438. S2CID 13410244.
- Matta A, Siu KW, Ralhan R (May 2012). "14-3-3 zeta as novel molecular target for cancer therapy". Expert Opinion on Therapeutic Targets. 16 (5): 515–23. doi:10.1517/14728222.2012.668185. PMID 22512284. S2CID 38941816.
- Joo Y, Schumacher B, Landrieu I, Bartel M, Smet-Nocca C, Jang A, et al. (October 2015). "Involvement of 14-3-3 in tubulin instability and impaired axon development is mediated by Tau" (PDF). FASEB Journal. 29 (10): 4133–44. doi:10.1096/fj.14-265009. PMID 26103986. S2CID 32696302.
- Jérôme M, Paudel HK (September 2014). "14-3-3ζ regulates nuclear trafficking of protein phosphatase 1α (PP1α) in HEK-293 cells". Archives of Biochemistry and Biophysics. 558: 28–35. doi:10.1016/j.abb.2014.06.012. PMID 24956593.
- Killoran RC, Fan J, Yang D, Shilton BH, Choy WY (2015). "Structural Analysis of the 14-3-3ζ/Chibby Interaction Involved in Wnt/β-Catenin Signaling". PLOS ONE. 10 (4): e0123934. Bibcode:2015PLoSO..1023934K. doi:10.1371/journal.pone.0123934. PMC 4409382. PMID 25909186.
- Qureshi HY, Li T, MacDonald R, Cho CM, Leclerc N, Paudel HK (September 2013). "Interaction of 14-3-3ζ with microtubule-associated protein tau within Alzheimer's disease neurofibrillary tangles". Biochemistry. 52 (37): 6445–55. doi:10.1021/bi400442d. PMID 23962087.
- Fang D, Hawke D, Zheng Y, Xia Y, Meisenhelder J, Nika H, et al. (April 2007). "Phosphorylation of beta-catenin by AKT promotes beta-catenin transcriptional activity". The Journal of Biological Chemistry. 282 (15): 11221–9. doi:10.1074/jbc.M611871200. PMC 1850976. PMID 17287208.
- Lee JJ, Lee JS, Cui MN, Yun HH, Kim HY, Lee SH, Lee JH (November 2014). "BIS targeting induces cellular senescence through the regulation of 14-3-3 zeta/STAT3/SKP2/p27 in glioblastoma cells". Cell Death & Disease. 5 (11): e1537. doi:10.1038/cddis.2014.501. PMC 4260756. PMID 25412315.
- Zhang Y, Li Y, Lin C, Ding J, Liao G, Tang B (2014). "Aberrant upregulation of 14-3-3σ and EZH2 expression serves as an inferior prognostic biomarker for hepatocellular carcinoma". PLOS ONE. 9 (9): e107251. Bibcode:2014PLoSO...9j7251Z. doi:10.1371/journal.pone.0107251. PMC 4165773. PMID 25226601.
- Chakravarti R, Gupta K, Swain M, Willard B, Scholtz J, Svensson LG, et al. (July 2015). "14-3-3 in Thoracic Aortic Aneurysms: Identification of a Novel Autoantigen in Large Vessel Vasculitis". Arthritis & Rheumatology. 67 (7): 1913–21. doi:10.1002/art.39130. PMC 4624269. PMID 25917817.
- Qin J, Wang S, Wang P, Wang X, Ye H, Song C, et al. (May 2019). "Autoantibody against 14-3-3 zeta: a serological marker in detection of gastric cancer". Journal of Cancer Research and Clinical Oncology. 145 (5): 1253–1262. doi:10.1007/s00432-019-02884-5. PMID 30887154. S2CID 81980933.
- Liu M, Liu X, Ren P, Li J, Chai Y, Zheng SJ, et al. (May 2014). "A cancer-related protein 14-3-3ζ is a potential tumor-associated antigen in immunodiagnosis of hepatocellular carcinoma". Tumour Biology. 35 (5): 4247–56. doi:10.1007/s13277-013-1555-8. PMC 4096569. PMID 24390614.
- McGowan J, Peter C, Chattopadhyay S, Chakravarti R (2019). "14-3-3ζ-A Novel Immunogen Promotes Inflammatory Cytokine Production". Frontiers in Immunology. 10: 1553. doi:10.3389/fimmu.2019.01553. PMC 6667649. PMID 31396202.
- McGowan J, Peter C, Kim J, Popli S, Veerman B, Saul-McBeth J, et al. (October 2020). "14-3-3ζ-TRAF5 axis governs interleukin-17A signaling". Proceedings of the National Academy of Sciences of the United States of America. 117 (40): 25008–25017. doi:10.1073/pnas.2008214117. PMID 32968020. S2CID 221884385.
- Kerr JF, Wyllie AH, Currie AR (August 1972). "Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics". British Journal of Cancer. 26 (4): 239–57. doi:10.1038/bjc.1972.33. PMC 2008650. PMID 4561027.
- Noutsios GT, Ghattas P, Bennett S, Floros J (July 2015). "14-3-3 isoforms bind directly exon B of the 5'-UTR of human surfactant protein A2 mRNA". American Journal of Physiology. Lung Cellular and Molecular Physiology. 309 (2): L147-57. doi:10.1152/ajplung.00088.2015. PMC 4504974. PMID 26001776.
- Lewis J, Veldhuizen RA (1996). "Surfactant: current and potential therapeutic application in infants and adults". Journal of Aerosol Medicine. 9 (1): 143–54. doi:10.1089/jam.1996.9.143. PMID 10160204.
- Filoche M, Tai CF, Grotberg JB (July 2015). "Three-dimensional model of surfactant replacement therapy". Proceedings of the National Academy of Sciences of the United States of America. 112 (30): 9287–92. Bibcode:2015PNAS..112.9287F. doi:10.1073/pnas.1504025112. PMC 4522812. PMID 26170310.
- Powell DW, Rane MJ, Chen Q, Singh S, McLeish KR (June 2002). "Identification of 14-3-3zeta as a protein kinase B/Akt substrate". The Journal of Biological Chemistry. 277 (24): 21639–42. doi:10.1074/jbc.M203167200. PMID 11956222.
- Garcia-Guzman M, Dolfi F, Russello M, Vuori K (February 1999). "Cell adhesion regulates the interaction between the docking protein p130(Cas) and the 14-3-3 proteins". The Journal of Biological Chemistry. 274 (9): 5762–8. doi:10.1074/jbc.274.9.5762. PMID 10026197.
- Yang H, Masters SC, Wang H, Fu H (June 2001). "The proapoptotic protein Bad binds the amphipathic groove of 14-3-3zeta". Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology. 1547 (2): 313–9. doi:10.1016/s0167-4838(01)00202-3. PMID 11410287.
- Clark GJ, Drugan JK, Rossman KL, Carpenter JW, Rogers-Graham K, Fu H, et al. (August 1997). "14-3-3 zeta negatively regulates raf-1 activity by interactions with the Raf-1 cysteine-rich domain". The Journal of Biological Chemistry. 272 (34): 20990–3. doi:10.1074/jbc.272.34.20990. PMID 9261098.
- Tzivion G, Luo ZJ, Avruch J (September 2000). "Calyculin A-induced vimentin phosphorylation sequesters 14-3-3 and displaces other 14-3-3 partners in vivo". The Journal of Biological Chemistry. 275 (38): 29772–8. doi:10.1074/jbc.M001207200. PMID 10887173.
- Koyama S, Williams LT, Kikuchi A (July 1995). "Characterization of the interaction of Raf-1 with ras p21 or 14-3-3 protein in intact cells". FEBS Letters. 368 (2): 321–5. doi:10.1016/0014-5793(95)00686-4. PMID 7628630. S2CID 29625141.
- Van Der Hoeven PC, Van Der Wal JC, Ruurs P, Van Dijk MC, Van Blitterswijk J (January 2000). "14-3-3 isotypes facilitate coupling of protein kinase C-zeta to Raf-1: negative regulation by 14-3-3 phosphorylation". The Biochemical Journal. 345 Pt 2 (2): 297–306. doi:10.1042/0264-6021:3450297. PMC 1220759. PMID 10620507.
- Chow CW, Davis RJ (January 2000). "Integration of calcium and cyclic AMP signaling pathways by 14-3-3". Molecular and Cellular Biology. 20 (2): 702–12. doi:10.1128/MCB.20.2.702-712.2000. PMC 85175. PMID 10611249.
- Mils V, Baldin V, Goubin F, Pinta I, Papin C, Waye M, et al. (March 2000). "Specific interaction between 14-3-3 isoforms and the human CDC25B phosphatase". Oncogene. 19 (10): 1257–65. doi:10.1038/sj.onc.1203419. PMID 10713667.
- Calverley DC, Kavanagh TJ, Roth GJ (February 1998). "Human signaling protein 14-3-3zeta interacts with platelet glycoprotein Ib subunits Ibalpha and Ibbeta". Blood. 91 (4): 1295–303. doi:10.1182/blood.V91.4.1295. PMID 9454760.
- Feng S, Christodoulides N, Reséndiz JC, Berndt MC, Kroll MH (January 2000). "Cytoplasmic domains of GpIbalpha and GpIbbeta regulate 14-3-3zeta binding to GpIb/IX/V". Blood. 95 (2): 551–7. doi:10.1182/blood.V95.2.551. PMID 10627461.
- Du X, Fox JE, Pei S (March 1996). "Identification of a binding sequence for the 14-3-3 protein within the cytoplasmic domain of the adhesion receptor, platelet glycoprotein Ib alpha". The Journal of Biological Chemistry. 271 (13): 7362–7. doi:10.1074/jbc.271.13.7362. PMID 8631758.
- Du X, Harris SJ, Tetaz TJ, Ginsberg MH, Berndt MC (July 1994). "Association of a phospholipase A2 (14-3-3 protein) with the platelet glycoprotein Ib-IX complex". The Journal of Biological Chemistry. 269 (28): 18287–90. PMID 8034572.
- Prymakowska-Bosak M, Hock R, Catez F, Lim JH, Birger Y, Shirakawa H, et al. (October 2002). "Mitotic phosphorylation of chromosomal protein HMGN1 inhibits nuclear import and promotes interaction with 14.3.3 proteins". Molecular and Cellular Biology. 22 (19): 6809–19. doi:10.1128/mcb.22.19.6809-6819.2002. PMC 134047. PMID 12215538.
- Sliva D, Gu M, Zhu YX, Chen J, Tsai S, Du X, Yang YC (February 2000). "14-3-3zeta interacts with the alpha-chain of human interleukin 9 receptor". The Biochemical Journal. 345 Pt 3 (3): 741–7. doi:10.1042/0264-6021:3450741. PMC 1220812. PMID 10642536.
- Birkenfeld J, Betz H, Roth D (January 2003). "Identification of cofilin and LIM-domain-containing protein kinase 1 as novel interaction partners of 14-3-3 zeta". The Biochemical Journal. 369 (Pt 1): 45–54. doi:10.1042/BJ20021152. PMC 1223062. PMID 12323073.
- Waterman MJ, Stavridi ES, Waterman JL, Halazonetis TD (June 1998). "ATM-dependent activation of p53 involves dephosphorylation and association with 14-3-3 proteins". Nature Genetics. 19 (2): 175–8. doi:10.1038/542. PMID 9620776. S2CID 26600934.
- Gannon-Murakami L, Murakami K (June 2002). "Selective association of protein kinase C with 14-3-3 zeta in neuronally differentiated PC12 Cells. Stimulatory and inhibitory effect of 14-3-3 zeta in vivo". The Journal of Biological Chemistry. 277 (26): 23116–22. doi:10.1074/jbc.M201478200. PMID 11950841.
- Zemlickova E, Dubois T, Kerai P, Clokie S, Cronshaw AD, Wakefield RI, et al. (August 2003). "Centaurin-alpha(1) associates with and is phosphorylated by isoforms of protein kinase C". Biochemical and Biophysical Research Communications. 307 (3): 459–65. doi:10.1016/S0006-291X(03)01187-2. PMID 12893243.
- De Valck D, Heyninck K, Van Criekinge W, Vandenabeele P, Fiers W, Beyaert R (September 1997). "A20 inhibits NF-kappaB activation independently of binding to 14-3-3 proteins". Biochemical and Biophysical Research Communications. 238 (2): 590–4. doi:10.1006/bbrc.1997.7343. PMID 9299557.
- Vincenz C, Dixit VM (August 1996). "14-3-3 proteins associate with A20 in an isoform-specific manner and function both as chaperone and adapter molecules". The Journal of Biological Chemistry. 271 (33): 20029–34. doi:10.1074/jbc.271.33.20029. PMID 8702721.
- Nellist M, Goedbloed MA, de Winter C, Verhaaf B, Jankie A, Reuser AJ, et al. (October 2002). "Identification and characterization of the interaction between tuberin and 14-3-3zeta". The Journal of Biological Chemistry. 277 (42): 39417–24. doi:10.1074/jbc.M204802200. PMID 12176984.
- Hashiguchi M, Sobue K, Paudel HK (August 2000). "14-3-3zeta is an effector of tau protein phosphorylation". The Journal of Biological Chemistry. 275 (33): 25247–54. doi:10.1074/jbc.M003738200. PMID 10840038.
Further reading
- Kino T, Pavlakis GN (April 2004). "Partner molecules of accessory protein Vpr of the human immunodeficiency virus type 1". DNA and Cell Biology. 23 (4): 193–205. doi:10.1089/104454904773819789. PMID 15142377.
- Kino T, Chrousos GP (June 2004). "Human immunodeficiency virus type-1 accessory protein Vpr: a causative agent of the AIDS-related insulin resistance/lipodystrophy syndrome?". Annals of the New York Academy of Sciences. 1024 (1): 153–67. Bibcode:2004NYASA1024..153K. doi:10.1196/annals.1321.013. PMID 15265780. S2CID 23655886.
- Calinisan V, Gravem D, Chen RP, Brittin S, Mohandas N, Lecomte MC, Gascard P (May 2006). "New insights into potential functions for the protein 4.1 superfamily of proteins in kidney epithelium". Frontiers in Bioscience. 11: 1646–66. doi:10.2741/1911. PMID 16368544.
PDB gallery | |
|---|---|
|