Gobiconodonta
Gobiconodonta is an order of extinct mammals known from the Early Jurassic (such as Huasteconodon) to early Late Cretaceous. They are generally held to be part of Eutriconodonta.[1]
| Gobiconodonta | |
|---|---|
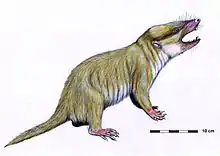 | |
| Life restoration of a Gobiconodon | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Clade: | Holotheria |
| Order: | †Gobiconodonta Averianov & Lopatin, 2011 |
| Families | |
Biology
Like many other non-therian mammals, gobiconodonts retained classical mammalian synapomorphies like epipubic bones (and likely the associated reproductive constrictions), venomous spurs and sprawling limbs. However, the forelimb and shoulder anatomy of at least some species like Jeholodens are similar to those of therian mammals, though the hindlimbs remain more conservative.[1] Gobiconodonts had a modern ear anatomy, the main difference from therians being that the ear ossicles were still somewhat connected to the jaw via the Meckel's cartilage.[2]
Some information on gobiconodont soft-tissues can be seen in Spinolestes, which was very well preserved, showing evidence of fur and internal organs. Spinolestes shows hair similar to that of modern mammals, with compound hair follicles with primary and secondary hair, even preserving traces of a pore infection. It also possesses a clear thoracic diaphragm like modern mammals, as well as spines, dermal scutes and an ossified Meckel's cartilage.[3]
The gobiconodont triconodont dentition has no analogue among living mammals, so comparisons are difficult. However, its clear that most if not all gobiconodonts were primarily carnivorous, given the presence of long, sharp canines and fang-like incisors,[note 1] premolars with trenchant main cusps that were well suited to grasp and pierce prey, strong development of the mandibular abductor musculature, bone crushing ability in at least some species and several other features.[1]
Gobiconodonts are often among the largest mammals in Mesozoic faunal assemblages, displaying a broad size range from small shrew-like insectivores to large forms like Repenomamus and Gobiconodon. They were among the first mammals to be specialised for vertebrate prey, and likely occupied the highest trophic levels among mammals in their faunal communities. Several forms like Gobiconodon and Repenomamus show evidence of scavenging, being among the few Mesozoic mammals to have significantly exploited that.[1]
At least in carnivorous niches, gobiconodonts were probably replaced by deltatheroidean metatherians, which are the dominant carnivorous mammals in Late Cretaceous faunal assemblages.[4] Competition between both groups is unattested, but in Asia the Early Cretaceous gobiconodontid diversity is replaced entirely by a deltatheroidean one, while in North America Nanocuris appears after the absence of Gobiconodon and other larger eutriconodonts.[5] Given that all insectivorous and carnivorous mammals groups suffered heavy losses during the mid-Cretaceous, it seems likely these metatherians simply occupied niches left after the extinction of eutriconodonts.[6]
At least Spinolestes had xenarthrous vertebrae and osseous scutes, convergent to those of modern xenarthrans and to a lesser extent the hero shrew.[3]
Uniquely among crown-group mammals, gobiconodontids replaced their molariform teeth by successors of similar complexity, while in other mammals less complex replacements are the norm.[7]
Classification
Cladogram after Averianov & Lopatin, 2011:[8]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Gobiconodonta was named by Averianov & Lopatin (2011); according to the authors it contained two families of mammals (gobiconodontids and klameliids)[8] that were traditionally assigned to Eutriconodonta.[9][1] The exact phylogenetic position of these families within Mammaliaformes is uncertain. Some analyses using only dental and mandibular characters (the first analysis of Gao et al., 2010,[10] Meng, Wang & Li, 2011,[11] Averianov & Lopatin, 2011[8]) found that gobiconodonts and eutriconodonts did not form a clade that wouldn't also include trechnotherians; however, some other analyses of dental and mandibular characters (Gaetano and Rougier, 2011,[12] 2012[13]) did recover gobiconodontids as eutriconodonts. The analysis conducted by Luo et al. (2007)[14] and the second analysis of Gao et al. (2010),[10] involving a broader range of Mesozoic mammaliaforms and more characters (including postcranial ones) recovered gobiconodontids as eutriconodonts as well. However, Gao et al. (2010) stressed that jeholodentids and gobiconodontids are the only eutriconodonts with known postcranial skeletons; according to the authors, it remains uncertain whether the results of their second analysis represent true phylogeny or are merely "a by-product of long branch attraction of jeholodentids and gobiconodontids".[10]
Most recent analysis recover them as eutriconodonts, closer to triconodontids and their immediate outgroups than to amphilestids:[15]
| Eutriconodonta |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Notes
- In Gobiconodon, the canine-like lower incisors outrightly replace the true canines, which are vestigial.[1]
References
- Zofia Kielan-Jaworowska; Richard L. Cifelli; Zhe-Xi Luo (2004). "Chapter 7: Eutriconodontans". Mammals from the Age of Dinosaurs: origins, evolution, and structure. New York: Columbia University Press. pp. 216–248. ISBN 0-231-11918-6.
- Meng Jing (2014). "Mesozoic mammals of China: implications for phylogeny and early evolution of mammals". Natl Sci Rev. 1 (4): 521–542. doi:10.1093/nsr/nwu070.
- Martin, Thomas; Marugán-Lobón, Jesús; Vullo, Romain; Martín-Abad, Hugo; Luo, Zhe-Xi; Buscalioni, Angela D. (2015). "A Cretaceous eutriconodont and integument evolution in early mammals". Nature. 526 (7573): 380–384. doi:10.1038/nature14905. PMID 26469049.
- Zofia Kielan-Jaworowska; Richard L. Cifelli; Zhe-Xi Luo (2004). "Chapter 12: Metaherians". Mammals from the Age of Dinosaurs: origins, evolution, and structure. New York: Columbia University Press. pp. 425–462. ISBN 0-231-11918-6.
- Rougier, G. W.; Davis, B. M.; Novacek, M. J. (2015). "A deltatheroidan mammal from the Upper Cretaceous Baynshiree Formation, eastern Mongolia". Cretaceous Research. 52: 167–177. doi:10.1016/j.cretres.2014.09.009.
- Grossnickle D. M., Polly P. D. (2013). "Mammal disparity decreases during the Cretaceous angiosperm radiation". Proceedings of the Royal Society B: Biological Sciences. 280: 20132110. doi:10.1098/rspb.2013.2110.
- https://onlinelibrary.wiley.com/doi/pdf/10.1002/spp2.1329
- A. O. Averianov; A. V. Lopatin (2011). "Phylogeny of Triconodonts and Symmetrodonts and the Origin of Extant Mammals". Doklady Biological Sciences. 436 (1): 32–35. doi:10.1134/s0012496611010042. PMID 21374009.
- Thomas Martin; Alexander O. Averianov (2006). "A previously unrecognized group of Middle Jurassic triconodontan mammals from Central Asia". Naturwissenschaften. 94 (1): 43–48. doi:10.1007/s00114-006-0155-5. PMID 17016686.
- Chun-Ling Gao; Gregory P. Wilson; Zhe-Xi Luo; A. Murat Maga; Qingjin Meng; Xuri Wang (2010). "A new mammal skull from the Lower Cretaceous of China with implications for the evolution of obtuse-angled molars and 'amphilestid' eutriconodonts". Proceedings of the Royal Society B: Biological Sciences. 277 (1679): 237–246. doi:10.1098/rspb.2009.1014. PMC 2842676. PMID 19726475.
- Jin Meng; Yuanqing Wang; Chuankui Li (2011). "Transitional mammalian middle ear from a new Cretaceous Jehol eutriconodont". Nature. 472 (7342): 181–185. doi:10.1038/nature09921. PMID 21490668.
- Leandro C. Gaetano & Guillermo W. Rougier (2011). "New materials of Argentoconodon fariasorum (Mammaliaformes, Triconodontidae) from the Jurassic of Argentina and its bearing on triconodont phylogeny". Journal of Vertebrate Paleontology. 31 (4): 829–843. doi:10.1080/02724634.2011.589877.
- Leandro C. Gaetano & Guillermo W. Rougier (2012). "First Amphilestid from South America: A Molariform from the Jurassic Cañadón Asfalto Formation, Patagonia, Argentina". Journal of Mammalian Evolution. 19 (4): 235–248. doi:10.1007/s10914-012-9194-1.
- Luo, Z.-X.; Chen, P.; Li, G.; Chen, M. (2007). "A new eutriconodont mammal and evolutionary development in early mammals". Nature. 446 (7133): 288–293. doi:10.1038/nature05627. PMID 17361176.
- Martin, Thomas; Marugán-Lobón, Jesús; Vullo, Romain; Martín-Abad, Hugo; Luo, Zhe-Xi; Buscalioni, Angela D. (2015). "A Cretaceous eutriconodont and integument evolution in early mammals". Nature. 526 (7573): 380–384. doi:10.1038/nature14905. PMID 26469049.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||