Piwi-interacting RNA
Piwi-interacting RNA (piRNA) is the largest class of small non-coding RNA molecules expressed in animal cells.[1][2] [3] piRNAs form RNA-protein complexes through interactions with piwi-subfamily Argonaute proteins. These piRNA complexes are mostly involved in the epigenetic and post-transcriptional silencing of transposable elements and other spurious or repeat-derived transcripts, but can also be involved in the regulation of other genetic elements in germ line cells.[4][5][6]
piRNAs are mostly created from loci that function as transposon traps which provide a kind of RNA-mediated adaptive immunity against transposon expansions and invasions.[7] They are distinct from microRNA (miRNA) in size (26–31 nucleotides as opposed to 21–24 nt), lack of sequence conservation, increased complexity, and independence of Dicer for biogenesis, at least in animals.[5][1][2] (Plant Dcl2 may play a role in rasi/piRNA biogenesis.)[8][9]
Double-stranded RNAs capable of silencing repeat elements, then known as repeat associated small interfering RNA (rasiRNA), were proposed in Drosophila in 2001.[10] By 2008, it was still unclear how piRNAs are generated, but potential methods had been suggested, and it was certain their biogenesis pathway is distinct from miRNA and siRNA, while rasiRNA is now considered a piRNA subspecies.[11]
Characteristics

piRNAs have been identified in both vertebrates and invertebrates, and although biogenesis and modes of action do vary somewhat between species, a number of features are conserved. piRNAs have no clear secondary structure motifs,[1][12] the length of a piRNA varies between species (from 21 to 31 nucleotides), and the bias for a 5’ uridine is common to piRNAs in both vertebrates and invertebrates. piRNAs in Caenorhabditis elegans have a 5’ monophosphate and a 3’ modification that acts to block either the 2’ or 3’ oxygen;[13] this has also been confirmed to exist in Drosophila melanogaster,[14] zebrafish,[15] mice,[16] and rats.[15] This 3’ modification is a 2’-O-methylation; the reason for this modification is not clear, but it has been suggested that it increases piRNA stability.[15][17]
More than 50,000 unique piRNA sequences have been discovered in mice and more than 13,000 in D. melanogaster.[18] It is thought that there are many hundreds of thousands of different piRNA species in mammals.[19]
History and loci
In the early 1980s, it was discovered that a single mutation in the fruit fly genome could specifically activate all copies of a retrovirus-like element called Gypsy in the female germline. The site of the mutations that made these Gypsies "dance" was thus called the flamenco locus. In 2001, Aravin et al. proposed that double-stranded (ds) RNA-mediated silencing is implicated in the control of retrotransposons in the germline and by 2003 the idea had emerged that vestiges of transposons might produce dsRNAs required for the silencing of "live" transposons.[10] Sequencing of the 200,000-bp flamenco locus was difficult, as it turned out to be packed with transposable element fragments (104 insertions of 42 different transposons, including multiple Gypsies), all facing the same direction. Indeed, piRNAs are all found in clusters throughout animal genomes; these clusters may contain as few as ten or many thousands of piRNAs matching different, phased transposon fragments. This led to the idea in 2007 that in germlines a pool of primary piRNAs is processed from long single-stranded transcripts encoded by piRNA clusters in the opposite orientation of the transposons, so that the piRNAs can anneal to and complement the transposon-encoded transcripts, thereby triggering their degradation. Any transposon landing in the correct orientation in such a cluster will make the individual more or less immune to that transposon, and such an advantageous mutation will spread quickly through the population. The original mutations in the flamenco locus inhibited the transcription of the master transcript, thereby deactivating this defense system.[7][20][1][21][22]
A historical example of invasion and Piwi response is known: the P-element transposon invaded a Drosophila melanogaster genome in the mid-20th century, and, through interbreeding, within decades all wild fruit flies worldwide (though not the reproductively isolated lab strains) contained the same P-element. Repression of further P-element activity, spreading near-simultaneously, appears to have occurred by the Piwi-interacting RNA pathway.[23]
piRNA clusters in genomes can now readily be detected via bioinformatics methods.[24] While D. melanogaster and vertebrate piRNAs have been located in areas lacking any protein-coding genes,[11][20] piRNAs in C. elegans have been identified amidst protein-coding genes.[13]
In mammals, piRNAs are found both in testes[25] and ovaries,[26] although they only seem to be required in males.[4] In invertebrates, piRNAs have been detected in both the male and female germlines.[15][19]
At the cellular level, piRNAs have been found within both the nucleus and cytoplasm, suggesting that piRNA pathways may function in both of these areas[11] and, therefore, may have multiple effects.[27]
Classification
There are at least three Argonaute (Ago) subfamilies that have been found in eukaryotes. Unlike the Ago subfamily which is present in animals, plants, and fission yeast, the Piwi subfamily has only been found in animals.[28] RasiRNA has been observed in Drosophila and some unicellular eukaryotes but its presence in mammals has not been determined, unlike piRNA which has been observed in many species of invertebrates and vertebrates including mammals;[29] however, since proteins which associate with rasiRNA are found in both vertebrates and invertebrates, it is possible that active rasiRNA exist and have yet to be observed in other animals. RasiRNAs have been observed in Schizosaccharomyces pombe, a species of yeast, as well in some plants, neither of which have been observed to contain the Piwi subfamily of Argonaute proteins.[8] It has been observed that both rasiRNA and piRNA are maternally linked, but more specifically it is the Piwi protein subfamily that is maternally linked and therefore leads to the observation that rasiRNA and piRNA are maternally linked.[30]
Biogenesis
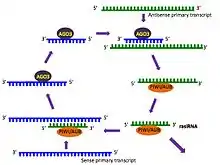
The biogenesis of piRNAs is not yet fully understood, although possible mechanisms have been proposed. piRNAs show a significant strand bias, that is, they are derived from one strand of DNA only,[1] and this may indicate that they are the product of long single stranded precursor molecules.[2] A primary processing pathway is suggested to be the only pathway used to produce pachytene piRNAs; in this mechanism, piRNA precursors are transcribed resulting in piRNAs with a tendency to target 5’ uridines.[31][32] Also proposed is a ‘Ping Pong’ mechanism wherein primary piRNAs recognise their complementary targets and cause the recruitment of piwi proteins. This results in the cleavage of the transcript at a point ten nucleotides from the 5’ end of the primary piRNA, producing the secondary piRNA.[32] These secondary piRNAs are targeted toward sequences that possess an adenine at the tenth position.[31] Since the piRNA involved in the ping pong cycle directs its attacks on transposon transcripts, the ping pong cycle acts only at the level of transcription.[22] One or both of these mechanisms may be acting in different species; C. elegans, for instance, does have piRNAs, but does not appear to use the ping pong mechanism at all.[19]
A significant number of piRNAs identified in zebrafish and D. melanogaster contain adenine at their tenth position,[11] and this has been interpreted as possible evidence of a conserved biosynthetic mechanism across species.[17] Ping-pong signatures have been identified in very primitive animals such as sponges and cnidarians, pointing to the existence of the ping-pong cycle already in the early branches of metazoans.[33]
Ping Pong
The piRNA Ping-Pong pathway was first proposed from studies in Drosophila where the piRNA associated with the two cytoplasmic Piwi proteins, Aubergine (Aub) and Argonaute-3 (Ago3) exhibited a high frequency of sequence complementarity over exactly 10 nucleotides at their 5′ ends.[32][34] This relationship is known as the "ping-pong signature" and is also observed in associated piRNA from Mili and Miwi2 proteins isolated from mouse testes. The proposed function of Ping-Pong in Drosophila or in mouse remains to be understood, but a leading hypothesis is that the interaction between Aub and Ago3 allows for a cyclic refinement of piRNA that are best suited to target active transposon sequences. Aub piRNA are primarily antisense to transposable element transcripts and are believed to be the main factor in targeting deleterious transcripts through complementarity. Conversely, Ago3 piRNA sequences are predominantly of sense orientation to transposable element transcripts and are derived from the product of Aub cleavage of transposon mRNA. As such, Ago3 piRNA lack the ability to target transposable element transcripts directly. Therefore, it was proposed that Ago3 piRNA guide the production of piRNA that are loaded into Aub by targeting newly exported piRNA cluster transcripts. Several lines of evidence support the effect of Ago3 on the production of Aub piRNA, in particular from examining the piRNA repertoire in Drosophila ovaries that are mutant for Ago3 and the Tudor-domain protein Kumo/Qin.[35][36]
The molecular mechanism that underpins Ping-Pong likely involves several piRNA pathway associated factors. Qin was reported to coordinate the loading of Ago3 with piRNA, in addition to interacting with both Aub and Ago3.[36] However, the Tudor protein krimper (A1ZAC4) was also shown to interact with both Aub and Ago3 through its Tudor domains while also binding itself through its N-terminal Krimper domain.[37] Specifically, Krimper interacts with Ago3 in its piRNA-unloaded state, while its interaction with Aub is dependent on the symmetrical dimethylation of arginine residues in the N-terminal region of Aub.[37][38] In Silkmoth germ cells, it was proposed that Vasa protein coordinates the Ping-Pong mechanism of Silkmoth Aub (Siwi) and Ago3.[39]
It is likely that the mechanism of Ping-Pong is primarily coordinated by Krimper but factors such as Kumo/Qin and Vasa, in addition to other factors have necessary functions in the Ping-Pong mechanism.
piRNA Phasing
The Drosophila piRNA pathway can be separated into two branches: the cytoplasmic branch consisting of Aub and Ago3 operating the Ping-Pong mechanism, and the nuclear branch, pertaining to the co-transcriptional silencing of genomic loci by Piwi in the nucleus. Through complementary strategies, two studies show that Aub and Ago3 target cleavage triggers the 'phased' loading of piRNA into Piwi.[40][41] Phasing begins with the targeting and cleavage of a complementary target by either Aub or Ago3 associated with a 'responder' piRNA. Once cleaved, the targeted transcript is then processed further by a mechanism believed to require the mitochondrial-associated endonuclease, Zucchini, which leads to the loading of Piwi protein with sequential fragments of the targeted transcript. In this way, the Aub or Ago3 'responder' piRNA sequence cleaves a complementary target that is then sliced at periodic intervals of approximately 27 nucleotides that are sequentially loaded into Piwi protein. Once loaded with piRNA, Piwi then enters the germ cell nucleus to co-transcriptionally silence nascent transcripts with complementarity to its piRNA guide.[42] It is currently unknown whether phasing occurs in other organisms.
Function
The wide variation in piRNA sequences and piwi function across species contributes to the difficulty in establishing the functionality of piRNAs.[43] However, like other small RNAs, piRNAs are thought to be involved in gene silencing,[1] specifically the silencing of transposons.[44] The majority of piRNAs are antisense to transposon sequences,[22] suggesting that transposons are targets of the piRNAs. In mammals, it appears that the activity of piRNAs in transposon silencing is most important during the development of the embryo,[31] and in both C. elegans and humans, piRNAs are necessary for spermatogenesis.[43]
RNA silencing
piRNA has a role in RNA silencing via the formation of an RNA-induced silencing complex (RISC). piRNAs interact with piwi proteins that are part of a family of proteins called the Argonautes. These are active in the testes of mammals and are required for germ-cell and stem-cell development in invertebrates. Three piwi subfamily proteins – MIWI, MIWI2, and MILI – have been found to be essential for spermatogenesis in mice. piRNAs direct the piwi proteins to their transposon targets.[31] A decrease or absence of PIWI gene expression is correlated with an increased expression of transposons.[11][31] Transposons have a high potential to cause deleterious effects on their hosts[21] and, in fact, mutations in piRNA pathways have been found to reduce fertility in D. melanogaster.[20] Further, it is thought that piRNA and endogenous small interfering RNA (endo-siRNA) may have comparable and even redundant functionality in transposon control in mammalian oocytes.[22]
piRNAs appear to affect particular methyltransferases that perform the methylations which are required to recognise and silence transposons,[31] but this relationship is not well understood.
Epigenetic effects
piRNAs can be transmitted maternally,[15] and based on research in D. melanogaster, piRNAs may be involved in maternally derived epigenetic effects.[20] The activity of specific piRNAs in the epigenetic process also requires interactions between piwi proteins and HP1a, as well as other factors.[18]
Accessory proteins of the piRNA pathway
Genetic screens examining fertility defects identified a number of proteins that are not Piwi-clade Argonautes, yet produce the same sterility phenotypes as Piwi mutants.
Drosophila Tudor domain proteins
Many factors required for the piRNA pathway in Drosophila contain Tudor domains that are known to bind symmetrically dimethylated arginine residues (sDMA) present in methylation motifs of Piwi proteins. Piwi proteins are symmetrically dimethylated by the PRMT5 methylosome complex, consisting of Valois (MEP50) and Capsulèen (dart5; PRMT5).[45][46]
- Tudor (Tud)
- Qin/Kumo
- Spindle-E (SpnE)
- Krimper
- Tejas (Tej)
- Vreteno (Vret)
- Papi
- Yb (fs(1)Yb)
- Brother of Yb (BoYB)
- Sister of Yb (SoYB)
Non-Tudor Drosophila piRNA pathway proteins
- Vasa
- Maelstrom (Mael)
Drosophila nuclear piRNA pathway proteins
- Rhino (HP1D)
- Deadlock
- Cutoff
- SetDB1 (Eggless)
- SuVar3–9
Investigation
Major advances in the study of piRNA have been achieved thanks to the use of next-generation sequencing techniques, such as Solexa, 454, and Illumina platform sequencing. These techniques allow analysis of highly complex and heterogeneous RNA populations like piRNAs. Due to their small size, expression and amplification of small RNAs can be challenging, so specialised PCR-based methods have been developed in response to this difficulty.[47][48] However, research has also revealed that a number of annotated piRNAs may be false positives; for instance, a majority of piRNAs that were expressed in somatic non-gonadal tissues were considered to derive from non-coding RNA fragments.[49]
References
- "Molecular Biology Select". Cell. 126 (2): 223–225. July 2006. doi:10.1016/j.cell.2006.07.012.
- Seto AG, Kingston RE, Lau NC (June 2007). "The coming of age for Piwi proteins". Molecular Cell. 26 (5): 603–609. doi:10.1016/j.molcel.2007.05.021. PMID 17560367.
- Monga I, Banerjee I (November 2019). "Computational Identification of piRNAs Using Features Based on RNA Sequence, Structure, Thermodynamic and Physicochemical Properties". Current Genomics. 20 (7): 508–518. doi:10.2174/1389202920666191129112705. PMC 7327968. PMID 32655289.
- Siomi MC, Sato K, Pezic D, Aravin AA (April 2011). "PIWI-interacting small RNAs: the vanguard of genome defence". Nature Reviews Molecular Cell Biology. 12 (4): 246–258. doi:10.1038/nrm3089. PMID 21427766. S2CID 5710813.
- Dorner S, Eulalio A, Huntzinger E, Izaurralde E (August 2007). "Delving into the diversity of silencing pathways. Symposium on MicroRNAs and siRNAs: biological functions and mechanisms". EMBO Reports. 8 (8): 723–729. doi:10.1038/sj.embor.7401015. PMC 1978081. PMID 17599087.
- Klattenhoff C, Bratu DP, McGinnis-Schultz N, Koppetsch BS, Cook HA, Theurkauf WE (January 2007). "Drosophila rasiRNA pathway mutations disrupt embryonic axis specification through activation of an ATR/Chk2 DNA damage response". Developmental Cell. 12 (1): 45–55. doi:10.1016/j.devcel.2006.12.001. PMID 17199040.
- Goriaux C, Théron E, Brasset E, Vaury C (2014). "History of the discovery of a master locus producing piRNAs: the flamenco/COM locus in Drosophila melanogaster". Frontiers in Genetics. 5: 257. doi:10.3389/fgene.2014.00257. PMC 4120762. PMID 25136352.
- Aravin A, Tuschl T (October 2005). "Identification and characterization of small RNAs involved in RNA silencing". FEBS Letters. 579 (26): 5830–5840. doi:10.1016/j.febslet.2005.08.009. PMID 16153643.
- Xie Z, Johansen LK, Gustafson AM, Kasschau KD, Lellis AD, Zilberman D, Jacobsen SE, Carrington JC (May 2004). "Genetic and functional diversification of small RNA pathways in plants". PLOS Biology. 2 (5): E104. doi:10.1371/journal.pbio.0020104. PMC 350667. PMID 15024409.
- Aravin AA, Naumova NM, Tulin AV, Vagin VV, Rozovsky YM, Gvozdev VA (July 2001). "Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D. melanogaster germline". Current Biology. 11 (13): 1017–1027. doi:10.1016/S0960-9822(01)00299-8. PMID 11470406. S2CID 14767819.
- Klattenhoff C, Theurkauf W (January 2008). "Biogenesis and germline functions of piRNAs". Development. 135 (1): 3–9. doi:10.1242/dev.006486. PMID 18032451.
- Carmen L, Michela B, Rosaria V, Gabriella M (2009). "Existence of snoRNA, microRNA, piRNA characteristics in a novel non-coding RNA: x-ncRNA and its biological implication in Homo sapiens". Journal of Bioinformatics and Sequence Analysis. 1 (2): 031–040.
- Ruby JG, Jan C, Player C, Axtell MJ, Lee W, Nusbaum C, Ge H, Bartel DP (December 2006). "Large-scale sequencing reveals 21U-RNAs and additional microRNAs and endogenous siRNAs in C. elegans". Cell. 127 (6): 1193–1207. doi:10.1016/j.cell.2006.10.040. PMID 17174894. S2CID 16838469.
- Vagin VV, Sigova A, Li C, Seitz H, Gvozdev V, Zamore PD (July 2006). "A distinct small RNA pathway silences selfish genetic elements in the germline". Science. 313 (5785): 320–324. Bibcode:2006Sci...313..320V. doi:10.1126/science.1129333. PMID 16809489. S2CID 40471466.
- Houwing S, Kamminga LM, Berezikov E, Cronembold D, Girard A, van den Elst H, et al. (April 2007). "A role for Piwi and piRNAs in germ cell maintenance and transposon silencing in Zebrafish". Cell. 129 (1): 69–82. doi:10.1016/j.cell.2007.03.026. hdl:11858/00-001M-0000-0012-E169-6. PMID 17418787. S2CID 13373509.
- Kirino Y, Mourelatos Z (April 2007). "Mouse Piwi-interacting RNAs are 2′-O-methylated at their 3′ termini". Nature Structural & Molecular Biology. 14 (4): 347–348. doi:10.1038/nsmb1218. PMID 17384647. S2CID 31193964.
- Faehnle CR, Joshua-Tor L (October 2007). "Argonautes confront new small RNAs". Current Opinion in Chemical Biology. 11 (5): 569–577. doi:10.1016/j.cbpa.2007.08.032. PMC 2077831. PMID 17928262.
- Lin H, Yin H, Beyret E, Findley S, Deng W (2008). "The role of the piRNA pathway in stem cell self-renewal". Developmental Biology. 319 (2): 479. doi:10.1016/j.ydbio.2008.05.048.
- Das PP, Bagijn MP, Goldstein LD, Woolford JR, Lehrbach NJ, Sapetschnig A, Buhecha HR, Gilchrist MJ, Howe KL, Stark R, Matthews N, Berezikov E, Ketting RF, Tavaré S, Miska EA (July 2008). "Piwi and piRNAs act upstream of an endogenous siRNA pathway to suppress Tc3 transposon mobility in the Caenorhabditis elegans germline". Molecular Cell. 31 (1): 79–90. doi:10.1016/j.molcel.2008.06.003. PMC 3353317. PMID 18571451.
- Brennecke J, Malone CD, Aravin AA, Sachidanandam R, Stark A, Hannon GJ (November 2008). "An epigenetic role for maternally inherited piRNAs in transposon silencing". Science. 322 (5906): 1387–1392. doi:10.1126/science.1165171. PMC 2805124. PMID 19039138.
- O'Donnell KA, Boeke JD (April 2007). "Mighty Piwis defend the germline against genome intruders". Cell. 129 (1): 37–44. doi:10.1016/j.cell.2007.03.028. PMC 4122227. PMID 17418784.
- Malone CD, Hannon GJ (February 2009). "Small RNAs as guardians of the genome". Cell. 136 (4): 656–668. doi:10.1016/j.cell.2009.01.045. PMC 2792755. PMID 19239887.
- Kelleher ES (August 2016). "Reexamining the P-Element Invasion of Drosophila melanogaster Through the Lens of piRNA Silencing". Genetics. 203 (4): 1513–1531. doi:10.1534/genetics.115.184119. PMC 4981261. PMID 27516614.
- Rosenkranz D, Zischler H (January 2012). "proTRAC—a software for probabilistic piRNA cluster detection, visualization and analysis". BMC Bioinformatics. 13 (5): 5. doi:10.1186/1471-2105-13-5. PMC 3293768. PMID 22233380.
- Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ, Kuramochi-Miyagawa S, Nakano T, Chien M, Russo JJ, Ju J, Sheridan R, Sander C, Zavolan M, Tuschl T (July 2006). "A novel class of small RNAs bind to MILI protein in mouse testes". Nature. 442 (7099): 203–207. doi:10.1038/nature04916. PMID 16751777. S2CID 4379895.
- Tam OH, Aravin AA, Stein P, Girard A, Murchison EP, Cheloufi S, Hodges E, Anger M, Sachidanandam R, Schultz RM, Hannon GJ (May 2008). "Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes". Nature. 453 (7194): 534–538. doi:10.1038/nature06904. PMC 2981145. PMID 18404147.
- Ruvkun G (July 2008). "Tiny RNA: Where do we come from? What are we? Where are we going?". Trends in Plant Science. 13 (7): 313–316. doi:10.1016/j.tplants.2008.05.005. PMID 18562240.
- Houwing S, Kamminga LM, Berezikov E, Cronembold D, Girard A, van den Elst H, Filippov DV, Blaser H, Raz E, Moens CB, Plasterk RH, Hannon GJ, Draper BW, Ketting RF (April 2007). "A role for Piwi and piRNAs in germ cell maintenance and transposon silencing in Zebrafish". Cell. 129 (1): 69–82. doi:10.1016/j.cell.2007.03.026. hdl:11858/00-001M-0000-0012-E169-6. PMID 17418787. S2CID 13373509.
- Girard A, Sachidanandam R, Hannon GJ, Carmell MA (July 2006). "A germline-specific class of small RNAs binds mammalian Piwi proteins". Nature. 442 (7099): 199–202. Bibcode:2006Natur.442..199G. doi:10.1038/nature04917. PMID 16751776. S2CID 3185036.
- Tomari Y, Du T, Haley B, Schwarz DS, Bennett R, Cook HA, Koppetsch BS, Theurkauf WE, Zamore PD (March 2004). "RISC assembly defects in the Drosophila RNAi mutant armitage". Cell. 116 (6): 831–841. doi:10.1016/S0092-8674(04)00218-1. PMID 15035985. S2CID 17588448.
- Aravin AA, Sachidanandam R, Bourc'his D, Schaefer C, Pezic D, Toth KF, Bestor T, Hannon GJ (September 2008). "A piRNA pathway primed by individual transposons is linked to de novo DNA methylation in mice". Molecular Cell. 31 (6): 785–799. doi:10.1016/j.molcel.2008.09.003. PMC 2730041. PMID 18922463.
- Brennecke J, Aravin AA, Stark A, Dus M, Kellis M, Sachidanandam R, Hannon GJ (March 2007). "Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila" (PDF). Cell. 128 (6): 1089–1103. doi:10.1016/j.cell.2007.01.043. PMID 17346786. S2CID 2246942.
- Grimson A, Srivastava M, Fahey B, Woodcroft BJ, Chiang HR, King N, Degnan BM, Rokhsar DS, Bartel DP (October 2008). "Early origins and evolution of microRNAs and Piwi-interacting RNAs in animals". Nature. 455 (7217): 1193–1197. doi:10.1038/nature07415. PMC 3837422. PMID 18830242.
- Gunawardane LS, Saito K, Nishida KM, Miyoshi K, Kawamura Y, Nagami T, Siomi H, Siomi MC (March 2007). "A slicer-mediated mechanism for repeat-associated siRNA 5′ end formation in Drosophila". Science. 315 (5818): 1587–1590. Bibcode:2007Sci...315.1587G. doi:10.1126/science.1140494. PMID 17322028. S2CID 11513777.
- Li C, Vagin VV, Lee S, Xu J, Ma S, Xi H, Seitz H, Horwich MD, Syrzycka M, Honda BM, Kittler EL, Zapp ML, Klattenhoff C, Schulz N, Theurkauf WE, Weng Z, Zamore PD (May 2009). "Collapse of germline piRNAs in the absence of Argonaute3 reveals somatic piRNAs in flies". Cell. 137 (3): 509–521. doi:10.1016/j.cell.2009.04.027. PMC 2768572. PMID 19395009.
- Zhang Z, Xu J, Koppetsch BS, Wang J, Tipping C, Ma S, Weng Z, Theurkauf WE, Zamore PD (November 2011). "Heterotypic piRNA Ping-Pong requires qin, a protein with both E3 ligase and Tudor domains". Molecular Cell. 44 (4): 572–584. doi:10.1016/j.molcel.2011.10.011. PMC 3236501. PMID 22099305.
- Webster A, Li S, Hur JK, Wachsmuth M, Bois JS, Perkins EM, Patel DJ, Aravin AA (August 2015). "Aub and Ago3 Are Recruited to Nuage through Two Mechanisms to Form a Ping-Pong Complex Assembled by Krimper". Molecular Cell. 59 (4): 564–575. doi:10.1016/j.molcel.2015.07.017. PMC 4545750. PMID 26295961.
- Sato K, Iwasaki YW, Shibuya A, Carninci P, Tsuchizawa Y, Ishizu H, Siomi MC, Siomi H (August 2015). "Krimper Enforces an Antisense Bias on piRNA Pools by Binding AGO3 in the Drosophila Germline". Molecular Cell. 59 (4): 553–563. doi:10.1016/j.molcel.2015.06.024. PMID 26212455.
- Xiol J, Spinelli P, Laussmann MA, Homolka D, Yang Z, Cora E, Couté Y, Conn S, Kadlec J, Sachidanandam R, Kaksonen M, Cusack S, Ephrussi A, Pillai RS (June 2014). "RNA clamping by Vasa assembles a piRNA amplifier complex on transposon transcripts". Cell. 157 (7): 1698–1711. doi:10.1016/j.cell.2014.05.018. PMID 24910301.
- Mohn F, Handler D, Brennecke J (May 2015). "Noncoding RNA. piRNA-guided slicing specifies transcripts for Zucchini-dependent, phased piRNA biogenesis". Science. 348 (6236): 812–817. doi:10.1126/science.aaa1039. PMC 4988486. PMID 25977553.
- Han BW, Wang W, Li C, Weng Z, Zamore PD (May 2015). "Noncoding RNA. piRNA-guided transposon cleavage initiates Zucchini-dependent, phased piRNA production". Science. 348 (6236): 817–821. doi:10.1126/science.aaa1264. PMC 4545291. PMID 25977554.
- Le Thomas A, Rogers AK, Webster A, Marinov GK, Liao SE, Perkins EM, Hur JK, Aravin AA, Tóth KF (February 2013). "Piwi induces piRNA-guided transcriptional silencing and establishment of a repressive chromatin state". Genes & Development. 27 (4): 390–399. doi:10.1101/gad.209841.112. PMC 3589556. PMID 23392610.
- Wang G, Reinke V (June 2008). "A C. elegans Piwi, PRG-1, regulates 21U-RNAs during spermatogenesis". Current Biology. 18 (12): 861–867. doi:10.1016/j.cub.2008.05.009. PMC 2494713. PMID 18501605.
- Ozata DM, Gainetdinov I, Zoch A, Phillip D, Zamore PD (2019). "PIWI-interacting RNAs: small RNAs with big functions". Nature Reviews Genetics. 20 (2): 89–108. doi:10.1038/s41576-018-0073-3. PMID 30446728.
- Kirino Y, Kim N, de Planell-Saguer M, Khandros E, Chiorean S, Klein PS, Rigoutsos I, Jongens TA, Mourelatos Z (May 2009). "Arginine methylation of Piwi proteins catalysed by dPRMT5 is required for Ago3 and Aub stability". Nat. Cell Biol. 11 (5): 652–8. doi:10.1038/ncb1872. PMC 2746449. PMID 19377467.
- Anne J, Mechler BM (May 2005). "Valois, a component of the nuage and pole plasm, is involved in assembly of these structures, and binds to Tudor and the methyltransferase Capsuléen". Development. 132 (9): 2167–77. doi:10.1242/dev.01809. PMID 15800004.
- Ro S, Park C, Jin J, Sanders KM, Yan W (December 2006). "A PCR-based method for detection and quantification of small RNAs". Biochemical and Biophysical Research Communications. 351 (3): 756–763. doi:10.1016/j.bbrc.2006.10.105. PMC 1934510. PMID 17084816.
- Tang F, Hayashi K, Kaneda M, Lao K, Surani MA (May 2008). "A sensitive multiplex assay for piRNA expression". Biochemical and Biophysical Research Communications. 369 (4): 1190–1194. doi:10.1016/j.bbrc.2008.03.035. PMC 3855189. PMID 18348866.
- Tosar JP, Rovira C, Cayota A (2018-01-22). "Non-coding RNA fragments account for the majority of annotated piRNAs expressed in somatic non-gonadal tissues". Communications Biology. 1 (1): 2. doi:10.1038/s42003-017-0001-7. PMC 6052916. PMID 30271890.
Further reading
- Lau NC, Seto AG, Kim J, Kuramochi-Miyagawa S, Nakano T, Bartel DP, Kingston RE (July 2006). "Characterization of the piRNA complex from rat testes". Science. 313 (5785): 363–367. doi:10.1126/science.1130164. PMID 16778019. S2CID 21150160.
- Kim VN (August 2006). "Small RNAs just got bigger: Piwi-interacting RNAs (piRNAs) in mammalian testes". Genes & Development. 20 (15): 1993–1997. doi:10.1101/gad.1456106. PMID 16882976.
- Girard A, Sachidanandam R, Hannon GJ, Carmell MA (July 2006). "A germline-specific class of small RNAs binds mammalian Piwi proteins". Nature. 442 (7099): 199–202. Bibcode:2006Natur.442..199G. doi:10.1038/nature04917. PMID 16751776. S2CID 3185036.
- Grivna ST, Beyret E, Wang Z, Lin H (July 2006). "A novel class of small RNAs in mouse spermatogenic cells". Genes & Development. 20 (13): 1709–1714. doi:10.1101/gad.1434406. PMC 1522066. PMID 16766680.
- Watanabe T, Takeda A, Tsukiyama T, Mise K, Okuno T, Sasaki H, Minami N, Imai H (July 2006). "Identification and characterization of two novel classes of small RNAs in the mouse germline: retrotransposon-derived siRNAs in oocytes and germline small RNAs in testes". Genes & Development. 20 (13): 1732–1743. doi:10.1101/gad.1425706. PMC 1522070. PMID 16766679.
- Carmell MA, Girard A, van de Kant HJ, Bourc'his D, Bestor TH, de Rooij DG, Hannon GJ (April 2007). "MIWI2 is essential for spermatogenesis and repression of transposons in the mouse male germline". Developmental Cell. 12 (4): 503–514. doi:10.1016/j.devcel.2007.03.001. PMID 17395546.
- Le Thomas A, Tóth KF, Aravin AA (January 2014). "To be or not to be a piRNA: genomic origin and processing of piRNAs". Genome Biology. 15 (1): 204. doi:10.1186/gb4154. PMC 4053809. PMID 24467990.
- Weick EM, Miska EA (September 2014). "piRNAs: from biogenesis to function". Development. 141 (18): 3458–3471. doi:10.1242/dev.094037. PMID 25183868.
External links
- PingPongPro – a software for finding ping-pong signatures and ping-pong cycle activity
- piRNA Bank – a web resource on classified and clustered piRNAs
- proTRAC – a software for probabilistic piRNA cluster detection, visualization, and analysis
- piRNA cluster – database
Types of nucleic acids | |||||||
|---|---|---|---|---|---|---|---|
| Constituents | |||||||
| Ribonucleic acids (coding, non-coding) |
| ||||||
| Deoxyribonucleic acids | |||||||
| Analogues | |||||||
| Cloning vectors | |||||||
| |||||||