Alu element
An Alu element is a short stretch of DNA originally characterized by the action of the Arthrobacter luteus (Alu) restriction endonuclease.[1] Alu elements are the most abundant transposable elements, containing over one million copies dispersed throughout the human genome.[2] Alu elements were thought to be selfish or parasitic DNA, because their sole known function is self reproduction. However they are likely to play a role in evolution and have been used as genetic markers.[3][4] They are derived from the small cytoplasmic 7SL RNA, a component of the signal recognition particle. Alu elements are highly conserved within primate genomes and originated in the genome of an ancestor of Supraprimates.[5]
Alu insertions have been implicated in several inherited human diseases and in various forms of cancer.
The study of Alu elements has also been important in elucidating human population genetics and the evolution of primates, including the evolution of humans.
Alu family
The Alu family is a family of repetitive elements in primate genomes, including the human genome.[6] Modern Alu elements are about 300 base pairs long and are therefore classified as short interspersed nuclear elements (SINEs) among the class of repetitive DNA elements. The typical structure is 5' - Part A - A5TACA6 - Part B - PolyA Tail - 3', where Part A and Part B (also known as "left arm" and "right arm") are similar nucleotide sequences. Expressed another way, it is believed modern Alu elements emerged from a head to tail fusion of two distinct FAMs (fossil antique monomers) over 100 million years ago, hence its dimeric structure of two similar, but distinct monomers (left and right arms) joined by an A-rich linker. Both monomers are thought to have evolved from 7SL, also known as SRP RNA.[7] The length of the polyA tail varies between Alu families.
There are over one million Alu elements interspersed throughout the human genome, and it is estimated that about 10.7% of the human genome consists of Alu sequences. However, less than 0.5% are polymorphic (i.e., occurring in more than one form or morph).[8] In 1988, Jerzy Jurka and Temple Smith discovered that Alu elements were split in two major subfamilies known as AluJ (named after Jurka) and AluS (named after Smith), and other Alu subfamilies were also independently discovered by several groups.[9] Later on, a sub-subfamily of AluS which included active Alu elements was given the separate name AluY. Dating back 65 million years, the AluJ lineage is the oldest and least active in the human genome. The younger AluS lineage is about 30 million years old and still contains some active elements. Finally, the AluY elements are the youngest of the three and have the greatest disposition to move along the human genome.[10] The discovery of Alu subfamilies led to the hypothesis of master/source genes, and provided the definitive link between transposable elements (active elements) and interspersed repetitive DNA (mutated copies of active elements).[11]
Related elements
B1 elements in rats and mice are similar to Alus in that they also evolved from 7SL RNA, but they only have one left monomer arm. 95% percent of human Alus are also found in chimpanzees, and 50% of B elements in mice are also found in rats. These elements are mostly found in introns and upstream regulatory elements of genes.[12]
The ancestral form of Alu and B1 is the fossil Alu monomer (FAM). Free-floating forms of the left and right arms exist, termed free left Alu monomers (FLAMs) and free right Alu monomers (FRAMs) respectively.[13] A notable FLAM in primates is the BC200 lncRNA.
Sequence features
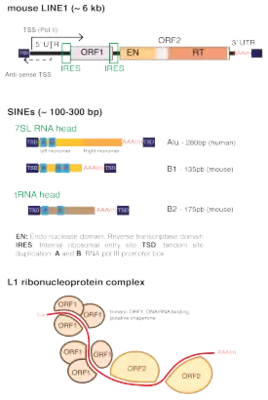
Two main promoter "boxes" are found in Alu: a 5' A box with the consensus TGGCTCACGCC, and a 3' B box with the consensus GWTCGAGAC (IUPAC nucleic acid notation). tRNAs, which are transcribed by RNA polymerase III, have a similar but stronger promoter structure.[14] Both boxes are located in the left arm.[7]
Alu elements contain four or fewer retinoic acid response element hexamer sites in its internal promoter, with the last one overlapping with the "B box".[15] In this 7SL (SRP) RNA example below, functional hexamers are underlined using a solid line, with the non-functional third hexamer denoted using a dotted line:
GCCGGGCGCGGTGGCGCGTGCCTGTAGTCCCagctACTCGGGAGGCTGAGGCTGGAGGATCGCTTGAGTCCAGGAGTTCTGGGCTGTAGTGCGCTATGCCGATCGGAATAGCCACTGCACTCCAGCCTGGGCAACATAGCGAGACCCCGTCTC.
The recognition sequence of the Alu I endonuclease is 5' ag/ct 3'; that is, the enzyme cuts the DNA segment between the guanine and cytosine residues (in lowercase above).[16]
Alu elements
Alu elements are responsible for regulation of tissue-specific genes. They are also involved in the transcription of nearby genes and can sometimes change the way a gene is expressed.[17]
Alu elements are retrotransposons and look like DNA copies made from RNA polymerase III-encoded RNAs. Alu elements do not encode for protein products. They are replicated as any other DNA sequence, but depend on LINE retrotransposons for generation of new elements.[18]
Alu element replication and mobilization begins by interactions with signal recognition particles (SRPs), which aid newly translated proteins to reach their final destinations.[19] Alu RNA forms a specific RNA:protein complex with a protein heterodimer consisting of SRP9 and SRP14.[19] SRP9/14 facilitates Alu's attachment to ribosomes that capture nascent L1 proteins. Thus, an Alu element can take control of the L1 protein's reverse transcriptase, ensuring that the Alu's RNA sequence gets copied into the genome rather than the L1's mRNA.[10]
Alu elements in primates form a fossil record that is relatively easy to decipher because Alu element insertion events have a characteristic signature that is both easy to read and faithfully recorded in the genome from generation to generation. The study of Alu Y elements (the more recently evolved) thus reveals details of ancestry because individuals will most likely only share a particular Alu element insertion if they have a common ancestor. This is because insertion of an Alu element occurs only 100 - 200 times per million years, and no known mechanism of deletion of one has been found. Therefore, individuals with an element likely descended from an ancestor with one—and vice versa, for those without. In genetics, the presence or lack thereof of a recently inserted Alu element may be a good property to consider when studying human evolution.[20]
Most human Alu element insertions can be found in the corresponding positions in the genomes of other primates, but about 7,000 Alu insertions are unique to humans.[21]
Impact of Alu in humans
Alu elements have been proposed to affect gene expression and been found to contain functional promoter regions for steroid hormone receptors.[15][22] Due to the abundant content of CpG dinucleotides found in Alu elements, these regions serve as a site of methylation, contributing to up to 30% of the methylation sites in the human genome.[23] Alu elements are also a common source of mutations in humans; however, such mutations are often confined to non-coding regions of pre-mRNA (introns), where they have little discernible impact on the bearer.[24] Mutations in the introns (or non-coding regions of RNA) have little or no effect on phenotype of an individual if the coding portion of individual's genome does not contain mutations. The Alu insertions that can be detrimental to the human body are inserted into coding regions (exons) or into mRNA after the process of splicing.[25]
However, the variation generated can be used in studies of the movement and ancestry of human populations,[26] and the mutagenic effect of Alu[27] and retrotransposons in general[28] has played a major role in the evolution of the human genome. There are also a number of cases where Alu insertions or deletions are associated with specific effects in humans:
Associations with human disease
Alu insertions are sometimes disruptive and can result in inherited disorders. However, most Alu variation acts as markers that segregate with the disease so the presence of a particular Alu allele does not mean that the carrier will definitely get the disease. The first report of Alu-mediated recombination causing a prevalent inherited predisposition to cancer was a 1995 report about hereditary nonpolyposis colorectal cancer.[29] In the human genome, the most recently active have been the 22 AluY and 6 AluS Transposon Element subfamilies due to their inherited activity to cause various cancers. Thus due to their major heritable damage it is important to understand the causes that affect their transpositional activity.[30]
The following human diseases have been linked with Alu insertions:[26][31]
- Alport syndrome
- Breast cancer
- chorioretinal degeneration
- Ewing's sarcoma
- Familial hypercholesterolemia
- Hemophilia
- Leigh syndrome
- mucopolysaccharidosis VII
- Neurofibromatosis
- Diabetes mellitus type II
And the following diseases have been associated with single-nucleotide DNA variations in Alu elements affecting transcription levels:[32]
Other Alu-associated human mutations
- The ACE gene, encoding angiotensin-converting enzyme, has 2 common variants, one with an Alu insertion (ACE-I) and one with the Alu deleted (ACE-D). This variation has been linked to changes in sporting ability: the presence of the Alu element is associated with better performance in endurance-oriented events (e.g. triathlons), whereas its absence is associated with strength- and power-oriented performance.[33]
- The opsin gene duplication which resulted in the re-gaining of trichromacy in Old World primates (including humans) is flanked by an Alu element,[34] implicating the role of Alu in the evolution of three colour vision.
References
- Schmid, Carl W; Deininger, Prescott L (1975). "Sequence organization of the human genome". Cell. 6 (3): 345–58. doi:10.1016/0092-8674(75)90184-1. PMID 1052772. S2CID 42804857.
- Szmulewicz, Martin N; Novick, Gabriel E; Herrera, Rene J (1998). "Effects of Alu insertions on gene function". Electrophoresis. 19 (8–9): 1260–4. doi:10.1002/elps.1150190806. PMID 9694261. S2CID 45917758.
- Kidwell, Margaret G; Lisch, Damon R (2001). "Perspective: Transposable Elements, Parasitic Dna, and Genome Evolution". Evolution. 55 (1): 1–24. doi:10.1554/0014-3820(2001)055[0001:ptepda]2.0.co;2. PMID 11263730.
- Pray, Leslie (2008). "Functions and Utility of Alu Jumping Genes". Scitable.com. Nature. Retrieved 26 June 2019.
- Kriegs, Jan Ole; Churakov, Gennady; Jurka, Jerzy; Brosius, Jürgen; Schmitz, Jürgen (2007). "Evolutionary history of 7SL RNA-derived SINEs in Supraprimates". Trends in Genetics. 23 (4): 158–61. doi:10.1016/j.tig.2007.02.002. PMID 17307271.
- Arcot, Santosh S.; Wang, Zhenyuan; Weber, James L.; Deininger, Prescott L.; Batzer, Mark A. (September 1995). "Alu Repeats: A Source for the Genesis of Primate Microsatellites". Genomics. 29 (1): 136–144. doi:10.1006/geno.1995.1224. ISSN 0888-7543. PMID 8530063.
- Häsler, Julien; Strub, Katharina (2006). "Alu elements as regulators of gene expression". Nucleic Acids Research. 34 (19): 5491–7. doi:10.1093/nar/gkl706. PMC 1636486. PMID 17020921.
- Roy-Engel, A. M; Carroll, M. L; Vogel, E; Garber, R. K; Nguyen, S. V; Salem, A. H; Batzer, M. A; Deininger, P. L (2001). "Alu insertion polymorphisms for the study of human genomic diversity". Genetics. 159 (1): 279–90. PMC 1461783. PMID 11560904.
- Jurka, J; Smith, T (1988). "A fundamental division in the Alu family of repeated sequences". Proceedings of the National Academy of Sciences. 85 (13): 4775–8. Bibcode:1988PNAS...85.4775J. doi:10.1073/pnas.85.13.4775. PMC 280518. PMID 3387438.
- Bennett, E. A; Keller, H; Mills, R. E; Schmidt, S; Moran, J. V; Weichenrieder, O; Devine, S. E (2008). "Active Alu retrotransposons in the human genome". Genome Research. 18 (12): 1875–83. doi:10.1101/gr.081737.108. PMC 2593586. PMID 18836035.
- Richard Shen, M; Batzer, Mark A; Deininger, Prescott L (1991). "Evolution of the master Alu gene(s)". Journal of Molecular Evolution. 33 (4): 311–20. Bibcode:1991JMolE..33..311R. doi:10.1007/bf02102862. PMID 1774786. S2CID 13091552.
- Tsirigos, Aristotelis; Rigoutsos, Isidore; Stormo, Gary D. (18 December 2009). "Alu and B1 Repeats Have Been Selectively Retained in the Upstream and Intronic Regions of Genes of Specific Functional Classes". PLOS Computational Biology. 5 (12): e1000610. Bibcode:2009PLSCB...5E0610T. doi:10.1371/journal.pcbi.1000610. PMC 2784220. PMID 20019790.
- Kojima, K. K. (16 August 2010). "Alu Monomer Revisited: Recent Generation of Alu Monomers". Molecular Biology and Evolution. 28 (1): 13–15. doi:10.1093/molbev/msq218. PMID 20713470.
- Conti, A; Carnevali, D; Bollati, V; Fustinoni, S; Pellegrini, M; Dieci, G (January 2015). "Identification of RNA polymerase III-transcribed Alu loci by computational screening of RNA-Seq data". Nucleic Acids Research. 43 (2): 817–35. doi:10.1093/nar/gku1361. PMC 4333407. PMID 25550429.
- Vansant, G; Reynolds, W. F (1995). "The consensus sequence of a major Alu subfamily contains a functional retinoic acid response element". Proceedings of the National Academy of Sciences. 92 (18): 8229–33. Bibcode:1995PNAS...92.8229V. doi:10.1073/pnas.92.18.8229. PMC 41130. PMID 7667273.
- Ullu E, Tschudi C (1984). "Alu sequences are processed 7SL RNA genes". Nature. 312 (5990): 171–2. Bibcode:1984Natur.312..171U. doi:10.1038/312171a0. PMID 6209580. S2CID 4328237.
- Britten, R. J (1996). "DNA sequence insertion and evolutionary variation in gene regulation". Proceedings of the National Academy of Sciences of the United States of America. 93 (18): 9374–7. Bibcode:1996PNAS...93.9374B. doi:10.1073/pnas.93.18.9374. PMC 38434. PMID 8790336.
- Kramerov, D; Vassetzky, N (2005). "Short Retroposons in Eukaryotic Genomes". International Review of Cytology. 247: 165–221. doi:10.1016/S0074-7696(05)47004-7. PMID 16344113.
- Weichenrieder, Oliver; Wild, Klemens; Strub, Katharina; Cusack, Stephen (2000). "Structure and assembly of the Alu domain of the mammalian signal recognition particle". Nature. 408 (6809): 167–73. Bibcode:2000Natur.408..167W. doi:10.1038/35041507. PMID 11089964. S2CID 4427070.
- Terreros, Maria C.; Alfonso-Sanchez, Miguel A.; Novick; Luis; Lacau; Lowery; Regueiro; Herrera (September 11, 2009). "Insights on human evolution: an analysis of Alu insertion polymorphisms". Journal of Human Genetics. 54 (10): 603–611. doi:10.1038/jhg.2009.86. PMID 19745832. S2CID 8153502.
- Chimpanzee Sequencing Analysis Consortium (2005). "Initial sequence of the chimpanzee genome and comparison with the human genome". Nature. 437 (7055): 69–87. Bibcode:2005Natur.437...69.. doi:10.1038/nature04072. PMID 16136131. S2CID 2638825.
- Norris, J; Fan, D; Aleman, C; Marks, J. R; Futreal, P. A; Wiseman, R. W; Iglehart, J. D; Deininger, P. L; McDonnell, D. P (1995). "Identification of a new subclass of Alu DNA repeats that can function as estrogen receptor-dependent transcriptional enhancers". The Journal of Biological Chemistry. 270 (39): 22777–82. doi:10.1074/jbc.270.39.22777. PMID 7559405. S2CID 45796017.
- Schmid, C. W (1998). "Does SINE evolution preclude Alu function?". Nucleic Acids Research. 26 (20): 4541–50. doi:10.1093/nar/26.20.4541. PMC 147893. PMID 9753719.
- Lander, Eric S; Linton, Lauren M; Birren, Bruce; Nusbaum, Chad; Zody, Michael C; Baldwin, Jennifer; Devon, Keri; Dewar, Ken; Doyle, Michael; Fitzhugh, William; Funke, Roel; Gage, Diane; Harris, Katrina; Heaford, Andrew; Howland, John; Kann, Lisa; Lehoczky, Jessica; Levine, Rosie; McEwan, Paul; McKernan, Kevin; Meldrim, James; Mesirov, Jill P; Miranda, Cher; Morris, William; Naylor, Jerome; Raymond, Christina; Rosetti, Mark; Santos, Ralph; Sheridan, Andrew; et al. (2001). "Initial sequencing and analysis of the human genome" (PDF). Nature. 409 (6822): 860–921. Bibcode:2001Natur.409..860L. doi:10.1038/35057062. PMID 11237011.
- Deininger, Prescott L; Batzer, Mark A (1999). "Alu Repeats and Human Disease". Molecular Genetics and Metabolism. 67 (3): 183–93. doi:10.1006/mgme.1999.2864. PMID 10381326. S2CID 15651921.
- Batzer, Mark A; Deininger, Prescott L (2002). "Alu Repeats and Human Genomic Diversity". Nature Reviews Genetics. 3 (5): 370–9. doi:10.1038/nrg798. PMID 11988762. S2CID 205486422.
- Shen, S; Lin, L; Cai, J. J; Jiang, P; Kenkel, E. J; Stroik, M. R; Sato, S; Davidson, B. L; Xing, Y (2011). "Widespread establishment and regulatory impact of Alu exons in human genes". Proceedings of the National Academy of Sciences. 108 (7): 2837–42. Bibcode:2011PNAS..108.2837S. doi:10.1073/pnas.1012834108. PMC 3041063. PMID 21282640.
- Cordaux, Richard; Batzer, Mark A (2009). "The impact of retrotransposons on human genome evolution". Nature Reviews Genetics. 10 (10): 691–703. doi:10.1038/nrg2640. PMC 2884099. PMID 19763152.
- Nyström-Lahti, Minna; Kristo, Paula; Nicolaides, Nicholas C; Chang, Sheng-Yung; Aaltonen, Lauri A; Moisio, Anu-Liisa; Järvinen, Heikki J; Mecklin, Jukka-Pekka; Kinzler, Kenneth W; Vogelstein, Bert; de la Chapelle, Albert; Peltomäki, Päivi (1995). "Founding mutations and Alu-mediated recombination in hereditary colon cancer". Nature Medicine. 1 (11): 1203–6. doi:10.1038/nm1195-1203. PMID 7584997. S2CID 39468812.
- Jin, Lingling; McQuillan, Ian; Li, Longhai (2017). "Computational identification of harmful mutation regions to the activity of transposable elements". BMC Genomics. 18 (Suppl 9): 862. doi:10.1186/s12864-017-4227-z. PMC 5773891. PMID 29219079.
- Deininger, Prescott (2011). "Alu elements: Know the SINEs". Genome Biology. 12 (12): 236. doi:10.1186/gb-2011-12-12-236. PMC 3334610. PMID 22204421.
- "SNP in the promoter region of the myeloperoxidase MPO gene". SNPedia. Archived from the original on 2010-05-21. Retrieved 2010-03-14.
- Puthucheary, Zudin; Skipworth, James RA; Rawal, Jai; Loosemore, Mike; Van Someren, Ken; Montgomery, Hugh E (2011). "The ACE Gene and Human Performance". Sports Medicine. 41 (6): 433–48. doi:10.2165/11588720-000000000-00000. PMID 21615186. S2CID 42531424.
- Dulai, K. S; von Dornum, M; Mollon, J. D; Hunt, D. M (1999). "The evolution of trichromatic color vision by opsin gene duplication in New World and Old World primates". Genome Research. 9 (7): 629–38. doi:10.1101/gr.9.7.629 (inactive 2021-01-11). PMID 10413401.CS1 maint: DOI inactive as of January 2021 (link)
External links
- Alu+Repetitive+Sequences at the US National Library of Medicine Medical Subject Headings (MeSH)
- "NCBI Genbank DNA encoding 7SL RNA". 2018-05-12. Cite journal requires
|journal=(help)
