Bird flight
Bird flight is the primary mode of locomotion used by most bird species in which birds take off and fly. Flight assists birds with feeding, breeding, avoiding predators, and migrating.

Bird flight is one of the most complex forms of locomotion in the animal kingdom. Each facet of this type of motion, including hovering, taking off, and landing, involves many complex movements. As different bird species adapted over millions of years through evolution for specific environments, prey, predators, and other needs, they developed specializations in their wings, and acquired different forms of flight.
Various theories exist about how bird flight evolved, including flight from falling or gliding (the trees down hypothesis), from running or leaping (the ground up hypothesis), from wing-assisted incline running or from proavis (pouncing) behavior.
Basic mechanics of bird flight
Lift and drag
The fundamentals of bird flight are similar to those of aircraft, in which the aerodynamic forces sustaining flight are lift and drag. Lift force is produced by the action of air flow on the wing, which is an airfoil. The airfoil is shaped such that the air provides a net upward force on the wing, while the movement of air is directed downward. Additional net lift may come from airflow around the bird's body in some species, especially during intermittent flight while the wings are folded or semi-folded[1][2] (cf. lifting body).
Aerodynamic drag is the force opposite to the direction of motion, and hence the source of energy loss in flight. The drag force can be separated into two portions, lift-induced drag, which is the inherent cost of the wing producing lift (this energy ends up primarily in the wingtip vortices), and parasitic drag, including skin friction drag from the friction of air and body surfaces and form drag from the bird's frontal area. The streamlining of bird's body and wings reduces these forces.
Wings

The bird's forelimbs (the wings) are the key to flight. Each wing has a central vane to hit the wind, composed of three limb bones, the humerus, ulna and radius. The hand, or manus, which ancestrally was composed of five digits, is reduced to three digits (digit II, III and IV or I, II, III depending on the scheme followed[3]), which serves as an anchor for the primaries, one of two groups of flight feathers responsible for the wing's airfoil shape. The other set of flight feathers, behind the carpal joint on the ulna, are called the secondaries. The remaining feathers on the wing are known as coverts, of which there are three sets. The wing sometimes has vestigial claws. In most species, these are lost by the time the bird is adult (such as the highly visible ones used for active climbing by hoatzin chicks), but claws are retained into adulthood by the secretarybird, screamers, finfoots, ostriches, several swifts and numerous others, as a local trait, in a few specimens.
Albatrosses have locking mechanisms in the wing joints that reduce the strain on the muscles during soaring flight.[4]
Even within a species wing morphology may differ. For example, adult European Turtle Doves have been found to have longer but more rounded wings than juveniles – suggesting that juvenile wing morphology facilitates their first migrations, while selection for flight maneuverability is more important after the juveniles’ first molt.[5]
Female birds exposed to predators during ovulation produce chicks that grow their wings faster than chicks produced by predator-free females. Their wings are also longer. Both adaptations may make them better at avoiding avian predators.[6]
Wing shape
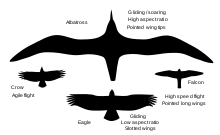
The shape of the wing is important in determining the flight capabilities of a bird. Different shapes correspond to different trade-offs between advantages such as speed, low energy use, and maneuverability. Two important parameters are the aspect ratio and wing loading. Aspect ratio is the ratio of wingspan to the mean of its chord (or the square of the wingspan divided by wing area). Wing loading is the ratio of weight to wing area.
Most kinds of bird wing can be grouped into four types, with some falling between two of these types. These types of wings are elliptical wings, high speed wings, high aspect ratio wings and soaring wings with slots.

Elliptical wings
Technically, elliptical wings are those having elliptical (that is quarter ellipses) meeting conformally at the tips. The early model Supermarine Spitfire is an example. Some birds have vaguely elliptical wings, including the albatross wing of high aspect ratio. Although the term is convenient, it might be more precise to refer to curving taper with fairly small radius at the tips. Many small birds have a low aspect ratio with elliptical character (when spread), allowing for tight maneuvering in confined spaces such as might be found in dense vegetation. As such they are common in forest raptors (such as Accipiter hawks), and many passerines, particularly non-migratory ones (migratory species have longer wings). They are also common in species that use a rapid take off to evade predators, such as pheasants and partridges.
High speed wings
High speed wings are short, pointed wings that when combined with a heavy wing loading and rapid wingbeats provide an energetically expensive, but high speed. This type of flight is used by the bird with the fastest wing speed, the peregrine falcon, as well as by most of the ducks. The same wing shape is used by the auks for a different purpose; auks use their wings to "fly" underwater.
The peregrine falcon has the highest recorded dive speed of 242 mph (389 km/h). The fastest straight, powered flight is the spine-tailed swift at 105 mph (170 km/h).

High aspect ratio wings
High aspect ratio wings, which usually have low wing loading and are far longer than they are wide, are used for slower flight. This may take the form of almost hovering (as used by kestrels, terns and nightjars) or in soaring and gliding flight, particularly the dynamic soaring used by seabirds, which takes advantage of wind speed variation at different altitudes (wind shear) above ocean waves to provide lift. Low speed flight is also important for birds that plunge-dive for fish.
Soaring wings with deep slots
These wings are favored by larger species of inland birds, such as eagles, vultures, pelicans, and storks. The slots at the end of the wings, between the primaries, reduce the induced drag and wingtip vortices by "capturing" the energy in air flowing from the lower to upper wing surface at the tips,[7] whilst the shorter size of the wings aids in takeoff (high aspect ratio wings require a long taxi to get airborne).[7]
Flight
Birds use three types of flight. They are distinguished by wing motion.
Gliding flight

When in gliding flight, the upward aerodynamic force is equal to the weight. In gliding flight, no propulsion is used; the energy to counteract the energy loss due to aerodynamic drag is either taken from the potential energy of the bird, resulting in a descending flight, or is replaced by rising air currents ("thermals"), referred to as soaring flight. For specialist soaring birds (obligate soarers), the decision to engage in flight are strongly related to atmospheric conditions that allow individuals to maximise flight-efficiency and minimise energetic costs.[8]
Flapping flight
When a bird flaps, as opposed to gliding, its wings continue to develop lift as before, but the lift is rotated forward to provide thrust, which counteracts drag and increases its speed, which has the effect of also increasing lift to counteract its weight, allowing it to maintain height or to climb. Flapping involves two stages: the down-stroke, which provides the majority of the thrust, and the up-stroke, which can also (depending on the bird's wings) provide some thrust. At each up-stroke the wing is slightly folded inwards to reduce the energetic cost of flapping-wing flight.[9] Birds change the angle of attack continuously within a flap, as well as with speed.[10]
Bounding flight
Small birds often fly long distances using a technique in which short bursts of flapping are alternated with intervals in which the wings are folded against the body. This is a flight pattern known as "bounding" or "flap-bounding" flight.[11] When the bird's wings are folded, its trajectory is primarily ballistic, with a small amount of body lift.[2] The flight pattern is believed to decrease the energy required by reducing the aerodynamic drag during the ballistic part of the trajectory,[12] and to increase the efficiency of muscle use.[13][14]
Hovering

Several bird species use hovering, with one family specialized for hovering – the hummingbirds.[15][16] True hovering occurs by generating lift through flapping alone, rather than by passage through the air, requiring considerable energy expenditure.[15][17] This usually confines the ability to smaller birds, but some larger birds, such as a kite[18] or osprey[19][20] can hover for a short period of time. Although not a true hover, some birds remain in a fixed position relative to the ground or water by flying into a headwind. Hummingbirds,[16][17] kestrels, terns and hawks use this wind hovering.
Most birds that hover have high aspect ratio wings that are suited to low speed flying. Hummingbirds are a unique exception – the most accomplished hoverers of all birds.[15] Hummingbird flight is different from other bird flight in that the wing is extended throughout the whole stroke, which is a symmetrical figure of eight,[21] with the wing producing lift on both the up- and down-stroke.[16][17] Hummingbirds beat their wings at some 43 times per second,[22] while others may be as high as 80 times per second.[23]
Take-off and landing


Take-off is one of the most energetically demanding aspects of flight, as the bird must generate enough airflow across the wing to create lift. Small birds do this with a simple upward jump. That doesn't work for larger birds, which must take a run up to generate sufficient airflow. Large birds take off by facing into the wind, or, if they can, by perching on a branch or cliff so they can just drop off into the air.
Landing is also a problem for large birds with high wing loads. This problem is dealt with in some species by aiming for a point below the intended landing area (such as a nest on a cliff) then pulling up beforehand. If timed correctly, the airspeed once the target is reached is virtually nil. Landing on water is simpler, and the larger waterfowl species prefer to do so whenever possible, landing into wind and using their feet as skids. To lose height rapidly prior to landing, some large birds such as geese indulge in a rapid alternating series of sideslips or even briefly turning upside down in a maneuver termed as whiffling.
Coordinated formation flight
A wide variety of birds fly together in a symmetric V-shaped or a J-shaped coordinated formation, also referred to as an "echelon", especially during long distance flight or migration. It is often assumed that birds resort to this pattern of formation flying in order to save energy and improve the aerodynamic efficiency.[24][25] The birds flying at the tips and at the front would interchange positions in a timely cyclical fashion to spread flight fatigue equally among the flock members.
The wingtips of the leading bird in an echelon create a pair of opposite rotating line vortices. The vortices trailing a bird have an underwash part behind the bird, and at the same time they have an upwash on the outside, that hypothetically could aid the flight of a trailing bird. In a 1970 study the authors claimed that each bird in a V formation of 25 members can achieve a reduction of induced drag and as a result increase their range by 71%.[26]
Studies of waldrapp ibis show that birds spatially coordinate the phase of wing flapping and show wingtip path coherence when flying in V positions, thus enabling them to maximally utilise the available energy of upwash over the entire flap cycle. In contrast, birds flying in a stream immediately behind another do not have wingtip coherence in their flight pattern and their flapping is out of phase, as compared to birds flying in V patterns, so as to avoid the detrimental effects of the downwash due to the leading bird's flight.[27]
Adaptations for flight

The most obvious adaptation to flight is the wing, but because flight is so energetically demanding birds have evolved several other adaptations to improve efficiency when flying. Birds' bodies are streamlined to help overcome air-resistance. Also, the bird skeleton is hollow to reduce weight, and many unnecessary bones have been lost (such as the bony tail of the early bird Archaeopteryx), along with the toothed jaw of early birds, which has been replaced with a lightweight beak. The skeleton's breastbone has also adapted into a large keel, suitable for the attachment of large, powerful flight muscles. The vanes of each feather have hooklets called barbules that zip the vanes of individual feathers together, giving the feathers the strength needed to hold the airfoil (these are often lost in flightless birds). The barbules maintain the shape and function of the feather. Each feather has a major (greater) side and a minor (lesser) side, meaning that the shaft or rachis does not run down the center of the feather. Rather it runs longitudinally of center with the lesser or minor side to the front and the greater or major side to the rear of the feather. This feather anatomy, during flight and flapping of the wings, causes a rotation of the feather in its follicle. The rotation occurs in the up motion of the wing. The greater side points down, letting air slip through the wing. This essentially breaks the integrity of the wing, allowing for a much easier movement in the up direction. The integrity of the wing is reestablished in the down movement, which allows for part of the lift inherent in bird wings. This function is most important in taking off or achieving lift at very low or slow speeds where the bird is reaching up and grabbing air and pulling itself up. At high speeds the air foil function of the wing provides most of the lift needed to stay in flight.
The large amounts of energy required for flight have led to the evolution of a unidirectional pulmonary system to provide the large quantities of oxygen required for their high respiratory rates. This high metabolic rate produces large quantities of radicals in the cells that can damage DNA and lead to tumours. Birds, however, do not suffer from an otherwise expected shortened lifespan as their cells have evolved a more efficient antioxidant system than those found in other animals.
Evolution of bird flight

Most paleontologists agree that birds evolved from small theropod dinosaurs, but the origin of bird flight is one of the oldest and most hotly contested debates in paleontology.[28] The four main hypotheses are:
- From the trees down, that birds' ancestors first glided down from trees and then acquired other modifications that enabled true powered flight.
- From the ground up, that birds' ancestors were small, fast predatory dinosaurs in which feathers developed for other reasons and then evolved further to provide first lift and then true powered flight.
- Wing-assisted incline running (WAIR), a version of "from the ground up" in which birds' wings originated from forelimb modifications that provided downforce, enabling the proto-birds to run up extremely steep slopes such as the trunks of trees.
- Pouncing proavis, which posits that flight evolved by modification from arboreal ambush tactics.
There has also been debate about whether the earliest known bird, Archaeopteryx, could fly. It appears that Archaeopteryx had the brain structures and inner-ear balance sensors that birds use to control their flight.[29] Archaeopteryx also had a wing feather arrangement like that of modern birds and similarly asymmetrical flight feathers on its wings and tail. But Archaeopteryx lacked the shoulder mechanism by which modern birds' wings produce swift, powerful upstrokes; this may mean that it and other early birds were incapable of flapping flight and could only glide.[30] The presence of most fossils in marine sediments in habitats devoid of vegetation has led to the hypothesis that they may have used their wings as aids to run across the water surface in the manner of the basilisk lizards.[31][32]
In March 2018, scientists reported that Archaeopteryx was likely capable of flight, but in a manner substantially different from that of modern birds.[33][34]
From the trees down
.jpg.webp)
This was the earliest hypothesis, encouraged by the examples of gliding vertebrates such as flying squirrels. It suggests that proto-birds like Archaeopteryx used their claws to clamber up trees and glided off from the tops.[35]
Some recent research undermines the "trees down" hypothesis by suggesting that the earliest birds and their immediate ancestors did not climb trees. Modern birds that forage in trees have much more curved toe-claws than those that forage on the ground. The toe-claws of Mesozoic birds and of closely related non-avian theropod dinosaurs are like those of modern ground-foraging birds.[36]
From the ground up
Feathers have been discovered in a variety of coelurosaurian dinosaurs (including the early tyrannosauroid Dilong).[37] Modern birds are classified as coelurosaurs by nearly all palaeontologists.[38] The original functions of feathers may have included thermal insulation and competitive displays. The most common version of the "from the ground up" hypothesis argues that bird's ancestors were small ground-running predators (rather like roadrunners) that used their forelimbs for balance while pursuing prey and that the forelimbs and feathers later evolved in ways that provided gliding and then powered flight.[39] Another "ground upwards" theory argues the evolution of flight was initially driven by competitive displays and fighting: displays required longer feathers and longer, stronger forelimbs; many modern birds use their wings as weapons, and downward blows have a similar action to that of flapping flight.[40] Many of the Archaeopteryx fossils come from marine sediments and it has been suggested that wings may have helped the birds run over water in the manner of the Jesus Christ lizard (common basilisk).[41]
Most recent attacks on the "from the ground up" hypothesis attempt to refute its assumption that birds are modified coelurosaurian dinosaurs. The strongest attacks are based on embryological analyses, which conclude that birds' wings are formed from digits 2, 3 and 4 (corresponding to the index, middle and ring fingers in humans; the first of a bird's 3 digits forms the alula, which they use to avoid stalling on low-speed flight, for example when landing); but the hands of coelurosaurs are formed by digits 1, 2 and 3 (thumb and first 2 fingers in humans).[42] However these embryological analyses were immediately challenged on the embryological grounds that the "hand" often develops differently in clades that have lost some digits in the course of their evolution, and therefore bird's hands do develop from digits 1, 2 and 3.[43][44][45]
Wing-assisted incline running
The wing-assisted incline running (WAIR) hypothesis was prompted by observation of young chukar chicks, and proposes that wings developed their aerodynamic functions as a result of the need to run quickly up very steep slopes such as tree trunks, for example to escape from predators. Note that in this scenario birds need downforce to give their feet increased grip.[46][47] But early birds, including Archaeopteryx, lacked the shoulder mechanism that modern birds' wings use to produce swift, powerful upstrokes. Since the downforce that WAIR requires is generated by upstrokes, it seems that early birds were incapable of WAIR.[30]
Pouncing proavis model
The proavis theory was first proposed by Garner, Taylor, and Thomas in 1999:
We propose that birds evolved from predators that specialized in ambush from elevated sites, using their raptorial hindlimbs in a leaping attack. Drag–based, and later lift-based, mechanisms evolved under selection for improved control of body position and locomotion during the aerial part of the attack. Selection for enhanced lift-based control led to improved lift coefficients, incidentally turning a pounce into a swoop as lift production increased. Selection for greater swooping range would finally lead to the origin of true flight.
The authors believed that this theory had four main virtues:
- It predicts the observed sequence of character acquisition in avian evolution.
- It predicts an Archaeopteryx-like animal, with a skeleton more or less identical to terrestrial theropods, with few adaptations to flapping, but very advanced aerodynamic asymmetrical feathers.
- It explains that primitive pouncers (perhaps like Microraptor) could coexist with more advanced fliers (like Confuciusornis or Sapeornis) since they did not compete for flying niches.
- It explains that the evolution of elongated rachis-bearing feathers began with simple forms that produced a benefit by increasing drag. Later, more refined feather shapes could begin to also provide lift.
Uses and loss of flight in modern birds
Birds use flight to obtain prey on the wing, for foraging, to commute to feeding grounds, and to migrate between the seasons. It is also used by some species to display during the breeding season and to reach safe isolated places for nesting.
Flight is more energetically expensive in larger birds, and many of the largest species fly by soaring and gliding (without flapping their wings) as much as possible. Many physiological adaptations have evolved that make flight more efficient.
Birds that settle on isolated oceanic islands that lack ground-based predators often lose the ability to fly. This illustrates both flight's importance in avoiding predators and its extreme demand for energy.
See also
Notes
- "Intermittent Flight Studies". Retrieved 6 March 2014.
- Tobalske, B; et al. "The intermittent flight of Zebra Finches: Unfixed gears and body lift". Retrieved 6 March 2014.
- Baumel JJ (1993) Handbook of Avian Anatomy: Nomina Anatomica Avium. 2nd Ed. Nuttall Ornithological Club. Cambridge, MA, USA
- Videler, JJ (2005) Avian Flight. Oxford University Press. ISBN 0-19-856603-4 pages 33-34
- Cabodevilla, X.; Moreno-Zarate, L.; Arroyo, B. (2018). "Differences in wing morphology between juvenile and adult European Turtle Doves Streptopelia turtur: implications for migration and predator escape". Ibis. 160 (2): 458–463. doi:10.1111/ibi.12564. hdl:10261/174622.
- Kaplan, Matt (25 March 2011). "Frightened birds grow longer wings". Nature. doi:10.1038/news.2011.187. Retrieved 27 March 2011.
- Tucker, Vance (July 1993). "Gliding Birds: Reduction of Induced Drag by Wing Tip Slots Between the Primary Feathers". Journal of Experimental Biology. 180: 285–310.
- Poessel, S. A.; Brandt, J.; Miller, T. A.; Katzner, T. E. (2018). "Meteorological and environmental variables affect flight behaviour and decision-making of an obligate soaring bird, the California Condor Gymnogyps californianus". Ibis. 160 (1): 36–53. doi:10.1111/ibi.12531.
- Parslew, B. (2012). Simulating Avian Wingbeats and Wakes, PhD Thesis
- Kristen E. Crandell & Bret W. Tobalske (2011). "Aerodynamics of tip-reversal upstroke in a revolving pigeon wing". The Journal of Experimental Biology. 214 (11): 1867–1873. doi:10.1242/jeb.051342. PMID 21562173.
- Bret W. Tobalske, Jason W. D. Hearn and Douglas R. Warrick, "Aerodynamics of intermittent bounds in flying birds", Exp. Fluids, 46, pp. 963–973 (2009), DOI 10.1007/s00348-009-0614-9 (accessed 2 August 2016)
- Brendan Body, Tips and observations of bird flight: "Further affects of air resistance on small birds", 2009 (accessed 2 August 2016)
- Tobalske, B.W., Peacock, W.L. & Dial, K.P. (1999). "Kinematics of flap-bounding flight in the Zebra Finch over a wide range of speeds" (PDF). The Journal of Experimental Biology. 202 (13): 1725–1739. PMID 10359676.CS1 maint: multiple names: authors list (link)
- Rayner J.M.V. (1985). "Bounding and undulating flight in birds". Journal of Theoretical Biology. 117 (1): 47–77. doi:10.1016/s0022-5193(85)80164-8.
- Ingersoll, Rivers; Haizmann, Lukas; Lentink, David (26 September 2018). "Biomechanics of hover performance in Neotropical hummingbirds versus bats". Science Advances. 4 (9): eaat2980. Bibcode:2018SciA....4.2980I. doi:10.1126/sciadv.aat2980. ISSN 2375-2548. PMC 6157961. PMID 30263957.
- Skandalis, Dimitri A.; Segre, Paolo S.; Bahlman, Joseph W.; Groom, Derrick J. E.; Welch, Kenneth C.; Witt, Christopher C.; McGuire, Jimmy A.; Dudley, Robert; Lentink, David; Altshuler, Douglas L. (19 October 2017). "The biomechanical origin of extreme wing allometry in hummingbirds". Nature Communications. 8 (1): 1047. Bibcode:2017NatCo...8.1047S. doi:10.1038/s41467-017-01223-x. ISSN 2041-1723. PMC 5715027. PMID 29051535.
- Ravi, S.; Crall, J. D.; McNeilly, L.; Gagliardi, S. F.; Biewener, A. A.; Combes, S. A. (12 March 2015). "Hummingbird flight stability and control in freestream turbulent winds". Journal of Experimental Biology. 218 (9): 1444–1452. doi:10.1242/jeb.114553. ISSN 0022-0949. PMID 25767146.
- Cascades Raptor Center (28 February 2012). "Cascades Raptor Center Show Behavior of the Year 2012". Retrieved 31 March 2018 – via YouTube.
- "Osprey General Information". www.newyorkwild.org. Retrieved 31 March 2018.
- Wild West Nature (4 April 2013). "Osprey hovers like a hummingbird hunting in Yellowstone National Park". Retrieved 31 March 2018 – via YouTube.
- Tobalske BW, Warrick DR, Clark CJ, Powers DR, Hedrick TL, Hyder GA, Biewener AA (2007). "Three-dimensional kinematics of hummingbird flight". J Exp Biol. 210 (13): 2368–82. doi:10.1242/jeb.005686. PMID 17575042.
- Hedrick, T. L.; Tobalske, B. W.; Ros, I. G.; Warrick, D. R.; Biewener, A. A. (14 December 2011). "Morphological and kinematic basis of the hummingbird flight stroke: scaling of flight muscle transmission ratio". Proceedings of the Royal Society B: Biological Sciences. 279 (1735): 1986–1992. doi:10.1098/rspb.2011.2238. ISSN 0962-8452. PMC 3311889. PMID 22171086.
- Gill V (30 July 2014). "Hummingbirds edge out helicopters in hover contest". BBC News. Retrieved 26 February 2019.
- Batt, Bruce (1 October 2007). "Why do migratory birds fly in a V-formation?". Scientific American. Retrieved 16 January 2014.
- Muijres, Florian T.; Dickinson, Michael H. (January 2014). "Fly with a little flap from your friends". Nature. 505 (7483): 295–296. doi:10.1038/505295a. ISSN 0028-0836. PMID 24429623. S2CID 4471158.
- Lissaman, P.B.S.; Shollenberger, Carl A. (22 May 1970). "Formation Flight of Birds". Science. 168 (3934): 1003–1005. Bibcode:1970Sci...168.1003L. doi:10.1126/science.168.3934.1003. PMID 5441020. S2CID 21251564.
- Portugal, Steven J.; Hubel, Tatjana Y.; Fritz, Johannes; Heese, Stefanie; Trobe, Daniela; Voelkl, Bernhard; Hailes, Stephen; Wilson, Alan M. & Usherwood, James R. (16 January 2014). "Upwash exploitation and downwash avoidance by flap phasing in ibis formation flight" (PDF). Nature. 505 (7483): 399–402. Bibcode:2014Natur.505..399P. doi:10.1038/nature12939. PMID 24429637. S2CID 205237135.
- Brush, A.H. (July 1998). "Taking Wing: Archaeopteryx and the Evolution of Bird Flight". The Auk. 115 (3): 806–808. doi:10.2307/4089435. JSTOR 4089435. Book review that provides a good, non-technical summary of the issues. The book is Shipman, P. (1999). Taking Wing: Archaeopteryx and the Evolution of Bird Flight. Simon & Schuster. ISBN 978-0-684-84965-2.
- Alonso, P.D.; Milner, A.C.; Ketcham, R.A.; Cokson, M.J & Rowe, T.B. (August 2004). "The avian nature of the brain and inner ear of Archaeopteryx". Nature. 430 (7000): 666–669. Bibcode:2004Natur.430..666A. doi:10.1038/nature02706. PMID 15295597. S2CID 4391019.
- Senter, P. (2006). "Scapular orientation in theropods and basal birds, and the origin of flapping flight" (Automatic PDF download). Acta Palaeontologica Polonica. 51 (2): 305–313.
- Videler, JJ (2005) Avian Flight. Oxford University Press. ISBN 0-19-856603-4 pages 98-117
- Videler, John (1 January 2005). "How Archaeopteryx could run over water". Archaeopteryx. 23. Retrieved 31 March 2018 – via ResearchGate.
- Voeten, Dennis F.A.E.; et al. (13 March 2018). "Wing bone geometry reveals active flight in Archaeopteryx". Nature Communications. 9 (923): 923. Bibcode:2018NatCo...9..923V. doi:10.1038/s41467-018-03296-8. PMC 5849612. PMID 29535376.
- Guarino, Ben (13 March 2018). "This feathery dinosaur probably flew, but not like any bird you know". The Washington Post. Retrieved 13 March 2018.
- Feduccia, A. (1999). The Origin and Evolution of Birds. Yale University Press. ISBN 978-0-300-07861-9. See also Feduccia, A. (February 1995). "Explosive Evolution in Tertiary Birds and Mammals". Science. 267 (5198): 637–638. Bibcode:1995Sci...267..637F. doi:10.1126/science.267.5198.637. PMID 17745839. S2CID 42829066.
- Glen, C.L. & Bennett, M.B. (November 2007). "Foraging modes of Mesozoic birds and non-avian theropods". Current Biology. 17 (21): R911–2. doi:10.1016/j.cub.2007.09.026. PMID 17983564. S2CID 535424. Archived from the original (abstract) on 8 December 2012.
- Prum, R. & Brush, A.H. (2002). "The evolutionary origin and diversification of feathers" (PDF). The Quarterly Review of Biology. 77 (3): 261–295. doi:10.1086/341993. PMID 12365352. S2CID 6344830. Archived from the original (PDF) on 15 October 2003. Retrieved 11 April 2019.
- Mayr G.; Pohl B.; Peters D.S. (2005). "A well-preserved Archaeopteryx specimen with theropod features". Science. 310 (5753): 1483–1486. Bibcode:2005Sci...310.1483M. doi:10.1126/science.1120331. PMID 16322455. S2CID 28611454.
- Burgers, P. & L. M. Chiappe (1999). "The wing of Archaeopteryx as a primary thrust generator". Nature. 399 (6731): 60–62. Bibcode:1999Natur.399...60B. doi:10.1038/19967. S2CID 4430686.
- Cowen, R. History of Life. Blackwell Science. ISBN 978-0-7266-0287-0.
- Videler, J.J. (2005). Avian Flight. Oxford: Oxford University Press. ISBN 978-0-19-856603-8.
- Burke, A.C. & Feduccia, A. (1997). "Developmental Patterns and the Identification of Homologies in the Avian Hand". Science. 278 (5338): 666–668. Bibcode:1997Sci...278..666B. doi:10.1126/science.278.5338.666. Summarized at "Embryo Studies Show Dinosaurs Could Not Have Given Rise To Modern Birds". ScienceDaily. October 1997.
- Chatterjee, S. (April 1998). "Counting the Fingers of Birds and Dinosaurs". Science. 280 (5362): 355a–355. Bibcode:1998Sci...280..355C. doi:10.1126/science.280.5362.355a.
- Vargas, A.O.; Fallon, J.F. (October 2004). "Birds have dinosaur wings: The molecular evidence" (abstract). Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 304B (1): 86–90. doi:10.1002/jez.b.21023. PMID 15515040.
- Pennisi, E. (January 2005). "Bird Wings Really Are Like Dinosaurs' Hands" (PDF). Science. 307 (5707): 194–195. doi:10.1126/science.307.5707.194b. PMID 15653478. S2CID 82490156. Archived from the original (PDF) on 27 July 2011.
- Dial, K.P. (2003). "Wing-Assisted Incline Running and the Evolution of Flight". Science. 299 (5605): 402–404. Bibcode:2003Sci...299..402D. doi:10.1126/science.1078237. PMID 12532020. S2CID 40712093. Summarized in Morelle, Rebecca (24 January 2008). "Secrets of bird flight revealed". Scientists believe they could be a step closer to solving the mystery of how the first birds took to the air. BBC News. Retrieved 25 January 2008.
- Bundle, M.W & Dial, K.P. (2003). "Mechanics of wing-assisted incline running (WAIR)" (PDF). The Journal of Experimental Biology. 206 (Pt 24): 4553–4564. doi:10.1242/jeb.00673. PMID 14610039. S2CID 6323207.
References
- Alexander, David E. Nature's Flyers: Birds, Insects, and the Biomechanics of Flight. 2002(hardcover) and 2004(paperback). Baltimore: The Johns Hopkins University Press. ISBN 0-8018-6756-8(hardcover) and 0801880599(paperback).
- Brooke, Michael and Tim Birkhead (editors). The Cambridge Encyclopedia of Ornithology. 1991. Cambridge: Cambridge University Press. ISBN 0-521-36205-9.
- Burton, Robert. Bird Flight. Facts on File, 1990
- Campbell, Bruce, and Elizabeth Lack (editors). A Dictionary of Birds. 1985. Calton: T&A D Poyse. ISBN 0-85661-039-9.
- Cornell Laboratory of Ornithology handbook of bird biology. 2004. Princeton University Press. ISBN 0-938027-62-X. (hardcover)
- Del Hoyo, Josep, et al. Handbook of Birds of the World Vol 1. 1992. Barcelona: Lynx Edicions, ISBN 84-87334-10-5.
- Wilson, Barry (editor). Readings from Scientific American, Birds. 1980. San Francisco: WH Freeman. ISBN 0-7167-1206-7.
- Attenborough, D.1998. The Life of Birds. Chapter 2. BBC Books. ISBN 0563-38792-0.
External links
| Wikimedia Commons has media related to Bird flight. |
- 'Flight in Birds and Aeroplanes' by Evolutionary Biologist John Maynard Smith Freeview video provided by the Vega Science Trust
- Beautiful Birds in Flight - slideshow by Life magazine
- 'Pigeon Take off in slow motion' YouTube video
- 'Bird Flight I' Eastern Kentucky University ornithology course site, with pictures, text and videos.