Djupadal Formation
The Djupadal Formation is a geologic formation in Skåne County, southern Sweden. It is Early (Pliensbachian to Toarcian) in age. It recovers at various locations throughout Central Skane, know by the discovery of basalt tuff layers, including Sandåkra, Korsaröd and Djupadal. An original analisis of the location of Korsaröd led to a Toarcian-Aalenian age.[3] Aspect that was recovered on various papers on the 80-90s.[1][4] But was dismissed on 2016, when a series of Palynogical samples recovered a Late Pliensbachian and probably Lower Toarcian age for the Korsaröd Outcrop.[5] The formation was deposited in the Central Skane region, linked to the late early Jurassic Volcanism. The Korsaröd member includes a volcanic-derived Lagerstatten with exceptional fern finds.[5] Thanks to the data provided by the fossilized wood rings was found that the location of Korsaröd hosted a middle-latitude Mediterranean-type biome in the late Early Jurassic, with low rainfall ratio, delayed to seasonal events. Superimposed on this climate were the effects of a local active Strombolian Volcanism and hydrothermal activity.[5]
| Djupadal Formation Stratigraphic range: Pliensbachian-Toarcian Boundary ~183 Ma | |
|---|---|
 Reconstruction of the environment at Korsaröd, based mostly on modern Rotorua | |
| Type | Formation |
| Underlies | Cuaternary Sediments |
| Overlies | Höör Sandstone Formation, Brandsberga and Kolleberga erratics and probably the Sapropel at Sandåkra |
| Thickness | Up to 60 m (200 ft)[1] |
| Lithology | |
| Primary | Basalt Tuff, Veined Gneiss[1] |
| Other | Sandstone, Clay and Conglomerate |
| Location | |
| Coordinates | 55.98°N 13.63°E |
| Region | Central Skåne County |
| Country | |
| Type section | |
| Named for | Djupadalsmölla, Ljungbyhed |
| Named by | Carita Augustsson[2] |
 Djupadal Formation (Sweden) Korsaröd Lagerstätten Location | |
Description

Djupadalsmölla, the original location has been known due to its volcanic Tuff and other volcanoclastic-derived facies.[2] It was originally described in 1826 as the local basalts where identified as coming from ancient volcanic eruptions.[6] The exposure of Anneklev, located near Höör was where the first volcanic neck was discovered, with other volcanic remnants mentioned on Jällabjär and Rallate along with the tuff at Djupadalsmölla.[6] These deposits were described as volcanic rocks composed mostly by tuff, that includes basaltic bombs (composed by Pyroxene and pseudomorphs from this, along with Olivine on adjacent pyroclastics), accidental lithics and occasional wood from conifers, in the form of small pieces to large logs.[7][8][9] At Djupadal the Rönne River has cut a 20 m deep valley mainly in the Precambrian basement, where Lower Jurassic strata (including volcanic tuff) form part of the southern valley side, and also occur in the valley northwestwards, indicating that the valley is partly an exhumed sub-Jurassic depression.[10] The modern strata of the Valley is over a Late Weichselian melt-water channel and it also contains eroded fluvioglacial deposits.[11] A nearby roadcut shows kaolinized basement beneath the Toarcian sediments and a nearby boring has penetrated 44 m of kaolinized Gneiss.[10] The Valley Bottom exposes small ridges and cupolas (in gneiss) and tors (mainly in Amphibolite).[10] The Djupadalsmölla pyroclastic reaches almost 10 m high and 20 m wide, with pyroclastics appearing also through the west more than 100 m, along the valley of the Rönne River.[7] The strata is composed by mostly small Lapilli (around 30–50% of the content) and ash (-10 mm), with some samples being red in patches. Tuff concretions are recovered locally composed mostly of coarse ash, with rich amounts of Calcite and Wood pieces.[2] This composition indicates moderate explosivity on the genesis of the materials, relating the eruption products with short transport paths, as show little mechanical weathering, also corroborated by the thick layering and the low amount of basaltic bombs reported, while the correlation of wood and lapilli indicates a terrestrial deposition.[2] All together shows local Strombolian Volcanism, linked probably to a coeval rift, as recovered by the presence of more than 100 coeval volcanic necks in central Scania.[2]
On Snälleröd (65 m thick, 44 m saprolite) samples taken, compared with the ones from the bottom of the road cut at Djupadal at 1.5 km NNE of this last one showed massive, soft lumps of a white, fine-grained material lacking any visually detectable grains of primary minerals, where only chemical data showed that this material is highly depleted in Calcium, Potassium and Sodium with significant kaolinization. The kaolinized rock has Kaolinite content exceeding 85%.[10] The samples taken at the NNE of Djudapal shows angular, gravel-sized material contains less altered granules of gneiss, remnant Feldspar and Quartz on a fraction dominated by Smectite, unlike the previous kaolinite-dominated ones, altrougth this last one is also abundant.[10]
The Korsaröd Lagerstatten is located also on central Scania, and represents the best outcrop of the formation, leading to exquisitely preserved (with fossilized nuclei and chromosomes) specimens of ferns of the extant genus Osmundastrum.[12][13] This location was linked by Ulf Sivhed in 1984 with the Dajupadal Formation.[4] What was corroborated by recent studies.[12] It is also composed by volcaniclastic deposits, located at 380 m WNW of the nearest basaltic volcanic plug.[5] It is composed by mafic clasts agruped with agglomerates, oriented to this volcanic plug, coming probably from it or nearby ones.[5] It´s clasts are angular and poorly sorted, recovered on a series of layers whose timing is uncertain, as there is no probe if represent discrete episodes separated by intervals of non-deposition or is result of variations due to a high-energy depositional setting.[5] Like in the Dajupadalsmölla type deposit, there is a great abundance of ash/mud content of the deposit filled with chaotic distributed wood fossil, what leds to the interpretation that this was a lahar deposit.[5] This location has been compared with modern Rotorua, New Zealand, considered an analogue for the type of environment represented in southern Sweden at this time.[14]
Eneskogen, Bonnarp and Säte volcanic necks are the main coeval of the Formation.[15] While Bonnarp (5-6 m height and covers roughly 5,000 square meters, covered by Jurassic sediments) is calculated to have at least 185.4+4.6 Ma (Middle Pliensbachian), Säte (Comprise two basalt pipes, each roughly 6–10 m high and some 10,000 square meters in area) yielded 180.0+0.7 Ma and Eneskogen (A large hill covered by quaternary sediments. Some few boulders and basalt pillars were exposed) 182.1+0.6 Ma, both Lower Toarcian in age.[15] It recovers one of the tree major mesozoic volcanic events on Skåne.[16] Säte & Bonnarp volcanic strata is composed by Basanite, being the first Glassy Facies, and the second microcrystalline, while Eneskogen also microcrystalline, is dominated by Melanephelinite.[17]
Age
Tralau (1973) measured the age for the local deposits, stablishing that where typical Middle Late Triassic to Lias strata, with absent Middle Jurassic sediments, with the exception of the volcanic event ejecting the tuffs in Korsaröd, stating that they took place in the Toarcian.[3] Radiometric ages obtained by using K–Ar techniques scatter in a wide range between 171 and 179 Ma.[18][19][20] Following works on the 80s and 90s recovered also this original datation, putting this and the Dajupadalsmölla outcrop on the Toarcian-Aalenian boundary, as example of latest lower jurassic volcanism on the region.[1][4] The Palynological studies on the 2014 tougth, changed the perspective of the age of the location, proposing a more fitting Late Pliensbachian.[12] In 2016 an in depth palynological study of the Korsaröd section led to stablish a Pliensbachian–early Toarcian(?) age, based on the high presence of the genus Perinopollenites elatoides (Pinales) and Eucommiidites troedsonii (Gnetales).[5] Other more recent works support that the outcrop is Lower Toarcian in age.[21] With a more recent work recovering it as 181.0-182.3 m.a., Lower Toarcian.[22] The formation overlies the Höor Sandstone Formation, and is time-equivalent with the Rydebäck and Katslösa members of the Rya Formation on NW Skane, the Röddinge Formation of the Vomb Trough and the Sorthat Formation of Denmark, with which it shares the abundance of Fern-derived material.[23] The formation also correlates with the Fjerritslev Formation of the Danish Basin,[24] and the Gassum Formation of the Øresund Basin.[25]
Pseudofungi
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Oomycetes Indeterminate |
|
|
A Pseudofungal Protist, Incertade sedis inside Oomycetes. Was recovered from the petiole of the holotype of Osmundastrum puchellum and interpreted as a peronosporomycete with parasitic or saprotrophic relation with this part of the plant.[26] |
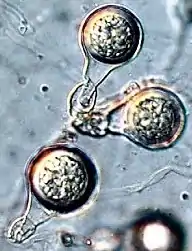 Extant Phytophthora, a Pseudofungal protist with sparsely bullate ornament, like the ones found Osmundastrum pulchellum | |
Fungi
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Fungi Indeterminate |
|
|
A Fungus, Incertade sedis inside Fungi. From the petiole and the root of Osmundastrum puchellum where recovered thread-like structures, identified as derived from a pathogenic or saprotrophic fungus invading necrotic tissues of the host plant.[26] |
_Gillet_845759.jpg.webp) Extant Microglossum, a saprotrophic fungus, whose Hypae are similar to the ones found Osmundastrum pulchellum | |
Palynology
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Botryococcus braunii |
|
|
A Freshwater Algae, type member of Botryococcaceae inside Chlorophyta. Goes from a 2%, being the most abundant algae on some samples to lack any presence on others.[5] |
 Extant Botryococcus | |
|
Pediastrum sp |
|
|
A Freshwater Algae, member of Hydrodictyaceae inside Chlorophyceae. The less abundant algae sampled locally.[5] |
 Extant Pediastrum | |
|
Lecaniella[5] |
Lecaniella sp. |
|
|
A Freshwater Algae, member of Zygnemataceae inside Charophyceae. On some samples is the only recovered algae, with a proportion of near a 3%.[5] |
|
|
|
|
A Miospore, member of Sphagnaceae inside Sphagnopsida. The major recovered Bryophyte spore, with less than 1.5% of the total samples.[5] |
 Extant Sphagnum, typical example of Sphagnaceae. Stereisporites probably come from a similar or a related Plant | |
|
Polycingulatisporites[5] |
Polycingulatisporites sp. |
|
|
A Miospore, related with the family Notothyladaceae inside Anthocerotopsida. Very scarce, with less than 1.1% on all the samples.[5] |
|
|
Neoraistrickia[5] |
Neoraistrickia sp. |
|
|
A Miospore, affinities with Selaginellaceae or Lycopodiaceae inside Lycopsida. The most abundant Lycopsid spore recovered locally, with near a 3.4% on some samples.[5] |
 Extant Selaginella, typical example of Selaginellaceae. Neoraistrickia probably come from a similar or a related Plant |
|
Retitriletes[5] |
|
|
|
A Miospore, affinities with Lycopodiaceae inside Lycopsida. Diverse, but less abundant, with a ratio of 1% by sample.[5] |
 Extant Austrolycopodium, typical example of Lycopodiaceae. Retitriletes probably come from a similar or a related Plant |
|
Calamospora[5] |
|
|
|
A Miospore, affinities with Sphenopsida inside Filicopsida. Rare, with a 4.3% on a single sample.[5] |
.JPG.webp) Extant Equisetum cone, the typical example of the Equisetopsida. Calamospora spores are pretty similar to the extant ones |
|
Conbaculatisporites[5] |
Conbaculatisporites mesozoicus |
|
|
A Miospore, incertade sedis inside Filicopsida. Scarce, with less than 2.3% on the samples.[5] |
|
|
Striatella seebergensis |
|
|
A Miospore, Incertade sedis inside Filicopsida. Very scarce, with less than 1.5% on all the samples.[5] |
||
|
Densoisporites[3] |
Densoisporites crassus |
|
|
A Miospore, affinities with Marsileaceae (specially Pilularia globulifera) inside Filicopsida. |
 Extant Pilularia, typical example of the Marsileaceae. Densoisporites may have come from a similar genus |
|
Lycopodiacidites[3] |
Lycopodiacidites rugulatus |
|
|
A Miospore, affinities with Ophioglossaceae inside Filicopsida. |
.jpg.webp) Extant Ophioglossum, typical example of the Ophioglossaceae. Lycopodiacidites may have come from a similar genus |
|
Marattisporites scabratus |
|
|
A Miospore, affinities with Marattiaceae inside Filicopsida. The second most abundant spore recovered on the location, with 3-14% on the samples.[5] |
 Extant Marattia, typical example of the Marattiaceae. Camptotriletes may have come from a similar genus | |
|
Osmundacidites wellmanii |
|
|
A Miospore, affinities with Osmundaceae inside Filicopsida. Can reach a 10% on some samples.[5] |
 Extant Osmunda, typical example of the Osmundaceae. Osmundacidites may have come from a similar genus | |
|
|
|
A Miospore, affinities with Osmundaceae inside Filicopsida. Very scarce, with less than 1-3.1% on all the samples.[5] |
 | |
|
Baculatisporites[3] |
|
|
|
A Miospore, affinities with Osmundaceae or Hymenophyllaceae inside Filicopsida. |
 Extant Hymenophyllum, typical example of the Hymenophyllaceae. Baculatisporites may have come from a similar genus |
|
Deltoidospora toralis |
|
|
A Miospore, related with Cyatheaceae, Dicksoniaceae and Schizaeaceae inside Filicopsida. Recovered on the petiole and the root of Osmundastrum puchellum.[26] |
_Domin_(11453662625).jpg.webp) Extant Cyathea, typical example of the Cyatheaceae. Deltoidospora may have come from a similar genus | |
|
Zebrasporites[3] |
|
|
|
A Miospore, related with Cyatheaceae inside Filicopsida. |
|
|
Cibotiumspora jurienensis |
|
|
A Miospore, related with Cyatheaceae inside Filicopsida. Recovered on the petiole and the root of Osmundastrum puchellum.[26] Present on various samples with ratios from 1-1.5%.[5] |
||
|
|
|
A Miospore, related with Cyatheaceae or Adiantaceae inside Filicopsida. Recovered on the petiole and the root of Osmundastrum puchellum.[26] It is the most common independent palynologycal residue recovered on Korsaröd, with Cyathidites spp. occuping a 21%.[5] Cyathidites minor almost certainly belong to well known Mesozoic species Coniopteris hymenophylloides and to other fossil cyatheaceous or dicksoniaceous ferns such as Eboracia lobifolia and Dicksonia mariopteri. |
_(14820801765).jpg.webp) | |
|
Contignisporites[3] |
Contignisporites problematicus |
|
|
A Miospore, related with Cibotiaceae inside Filicopsida. |
 Extant Cibotium, typical example of the Cibotiaceae. Contignisporites may have come from a similar genus |
|
|
|
A Pollen Grain, afinnities with Caytoniales inside Gymnospermopsida. Scarce, with around a 1% on all samples.[5] |
||
|
|
|
A Pollen Grain, Incertade sedis inside Pteridospermae. A. grandis reaches near a 10% on some samples.[5] |
||
|
|
|
A Pollen Grain, afinnities with Erdtmanithecales inside Spermatophytes. Moderately abundant, with around a 3-10% on all samples.[5] The Gymnosperms that produced Eucommiidites troedsonii pollen possibly dominated the understorey vegetation. Thick tectum, infratectum of small granules, indistinct or absent foot layer. Originally was thought to come from Angiosperms, latter reports suggest it come from arbustive Bennetites.[27] It was recently found to come from Eucommiitheca, member of the enigmatic Erdtmanithecales, reinterpreted as an unusual gymnosperm grain with a single distal colpus flanked by two subsidiary lateral colps.[28] Is very similar to the Pollen of the extant Ephedra and Welwitschia (mainly on the granular structure of the exine).[29] |
||
|
|
|
A Pollen Grain, incertade sedis inside Cycadopsida. Very rare, with less than 0.5% on all samples.[5] |
 Extant Encephalartos, typical example of the Cycas. Monosulcites Pollen is pretty similar to the extant ones of this genus | |
|
|
|
A Pollen Grain, incertade sedis inside Cycadopsida. Rare, with around a 0.7-1% on all samples.[5] |
 Extant Encephalartos, typical example of the Cycas. Chasmatosporites Pollen is pretty similar to the extant ones of this genus. Alternatively, it maybe come from Ginkos or Gnetales | |
|
|
|
A Pollen Grain, afinnities with Ginkgoales inside Ginkgophyta. Very rare, with less than 1% on all samples.[5] |
 Extant Ginkgo, only surviving example of the Ginkgoaceae. Ginkgoites Pollen is pretty similar to the extant ones of this genus | |
|
Quadraeculina[5] |
|
|
|
A Pollen Grain, incertade sedis inside Coniferophyta. Around a 1-3.9% on all the samples.[5] |
|
|
Rugaletes[5] |
|
|
|
A Pollen Grain, incertade sedis inside Coniferophyta. Around a 7.5% on all the samples.[5] |
|
|
Cerebropollenites[5] |
|
|
|
A Pollen Grain, affinities with Abietoideae inside Coniferophyta. Around 1-1.5% on all the samples.[5] Pollen From arbustive to arboreal plants, resembling the pollen of the modern genus Tsuga. The differences observed between Cerebropollenites and Tsuga are no greater than the differences observed between the pollen of the two Sections of Tsuga, Hesperopeuce and Micropeuce. |
 Extant Tsuga Cone, example of the Abietoideae. Cerebropollenites is similar to the pollen found on this genus |
|
Pinuspollenites[5] |
|
|
|
A Pollen Grain, affinities with Pinaceae inside Coniferophyta. Around a 1-4.5% on the samples.[5] |
 Extant Pinus cembra Cone, example of the Pinidae. Pinuspollenites is similar to the pollen found on this genus |
|
Cedripites[5] |
|
|
|
A Pollen Grain, affinities with Pinaceae inside Coniferophyta. Rare, with around a 1.5% on all samples.[5] |
|
|
|
|
A Pollen Grain, affinities with Cheirolepidiaceae inside Coniferophyta. Moderately abundant, with around a 1.5-9% on all samples.[5] |
||
|
Spheripollenites[5] |
|
|
|
A Pollen Grain, affinities with Cheirolepidiaceae inside Coniferophyta. Scarce, around a 1.5–3.9% on all the samples.[5] Abundant on the Lower Jurassic of NW Europe. Spheripollenites co-occurs on the coeval Sorthat Formation with cuticles of Dactyletrophyllum ramonensis, and after a test of relationships it was found a highly significant correlation may suggest that S. psilatus is produced by the conifer genus Dactyletrophyllum.[30] |
|
|
Parvisaccites[3] |
|
|
|
A Pollen Grain, affinities with Podocarpaceae and Cupressaceae inside Coniferophyta. |
.jpg.webp) Extant Podocarpus Cone, example of the Podocarpaceae. Parvisaccites is similar to the pollen found on this genus |
|
|
|
A Pollen Grain, affinities with Taxodiaceae and Cupressaceae inside Coniferophyta. The most abundant pollen recovered locally, with near a 30% on some samples.[5] |
 Extant Thuja Cone, example of the Cupressaceae. Perinopollenites is similar to the pollen found on this genus | |
|
Araucariacites[5] |
|
|
|
A Pollen Grain, afinnities with Araucariaceae inside Coniferophyta. Rare, with around a 1.5% on all samples.[5] |
 Extant Araucaria Cone, example of the Araucariaceae. Araucariacites is similar to the pollen found on this genus |

Megaflora
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Lycopsida[26] |
Lycopsida Indeterminate |
|
|
A Lycopsid, Incertade sedis inside Lycopsida. External exotic roots are preserved within detritus-filled cavities between the petiole bases of Osmundastrum pulchellum, with wall thickenings similar to the vasculature evident in ancient and modern herbaceous lycopsids.[26] |
 Extant Lycopodiella, a Lycopsid that grows it´s roots on floor ferns |
|
Osmundastrum pulchellum |
|
|
A Royal Fern, member of Osmundaceae inside Filicopsida. The most known fossil of the location, thanks to its exceptional fossilized Rhizome, that has preserved Nuclei and Chromosomes, a fine subcellular detail has rarely been documented in fossils.[12] It´s Rooted in DNA content was used to extrapolate relative genome, finding relationships with extant Osmundastrum cinnamomeum, and confirmed a monophyletic Osmunda.[12][31] Osmundastrum pulchellum is one of the earliest fossil Osmundastrum rhizomes known so far, and the first of its kind from the Mesozoic of Europe.[13] Its impressive preservation has lead to know even the biotic interactions with the Plant.[5] It also has recovered the only know case know to preserve the ongoing mitosis processes in plant cells via calcification from volcanic hydrothermal brine.[34] |
 Osmundastrum cinnamomeum, the most know extant relative of O. puchellum | |
|
Ptilophyllum[26] |
Ptilophyllum sp. |
|
|
A Bennetite, afinnities with Williamsoniaceae inside Bennettitales. This single impression of a bennettitalean leaf fragment found in a fine ash layer constituted the only foliar remains identified within the volcaniclastic deposit.[26] |
 Example of Ptilophyllum leaf |
Fossil Wood
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Protophyllocladoxylon[5] |
Protophyllocladoxylon sp. |
|
|
A Conifer, affinities with Cupressaceae inside Coniferophyta. The only diagnostic wood recovered locally, identified based on the possession of uniseriate rays with smooth walls, pointed oblique oopores, absence of axial parenchyma, and tracheid radial walls. It resembles the extant Thuja plicata. A few fossil wood specimens clearly represent portions of large trunks, with at least one fragment derived from a trunk of 1.68 m in diameter (with stimated c.5.3 m). Altrougth, most specimens represent lateral branches or even roots of small size (10 cm long and 5 cm wide).[5] It´s growth rings are distinct in all wood samples.[5] |
 Permineralized wood with a branch trace recovered from the volcaniclastic sediments at Korsaröd |
Arachnida
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Oribatida Indeterminate |
|
|
A Mite, Incertade sedis inside Oribatida. On the petiole of Osmundastrum puchellum excavations up to 715 μm in diameter are evident, filled with pellets that resemble the coprolites of Oribatid mites found also on Paleozoic and Mesozoic Woods.[26] |
 example of Oribatida mite | |
See also
- List of fossiliferous stratigraphic units in Sweden
- Kristianstad Basin
- Toarcian formations
- Rya Formation, Sweden
- Marne di Monte Serrone, Italy
- Calcare di Sogno, Italy
- Sachrang Formation, Austria
- Saubach Formation, Austria
- Posidonia Shale, Lagerstätte in Germany
- Ciechocinek Formation, Germany and Poland
- Krempachy Marl Formation, Poland and Slovakia
- Lava Formation, Lithuania
References
- Lidmar-Bergström, K., Olsson, S., & Olvmo, M. (1997). Palaeosurfaces and associated saprolites in southern Sweden. Geological Society, London, Special
- Augustsson, C. (2001). Lapilli tuff as evidence of Early Jurassic Strombolian-type volcanism in Scania, southern Sweden. GFF, 123(1), 23-28.
- Tralau, H.,1973 : En palynologisk åldersbestämning av v ulkanisk aktivitet i Skåne. -Fauna och Flora, 68, pp .12 1- 176. S tockholm.
- Sivhed, U. (1984). Litho-and biostratigraphy of the Upper Triassic-Middle Jurassic in Scania, southern Sweden (Vol. 78, No. 4). Sveriges geologiska undersökning.
- Vajda, V., Linderson, H., & McLoughlin, S. (2016). Disrupted vegetation as a response to Jurassic volcanism in southern Sweden. Geological Society, London, Special Publications, 434(1), 127-147.
- Kjellen, R., 1902: Bidrag till Sveriges endogena geografi. Geologiska Föreningens i Stockholm Förhandlingar 24, 193–220
- Eichstädt, F., 1883a: Om basalttuffen vid Djupadal i Skåne. Geologiska Föreningens i Stockholm Förhandlingar 6, 408–415.
- Eichstädt, F., 1883b: Ytterligare om basalttuffen vid Djupadal i Skåne. Geologiska Föreningens i Stockholm Förhandlingar 6, 774–783.
- Ahlberg, A., Sivhed, U. & Erlström, M., 2001: The Jurassic sediments in Skåne, southern Sweden. In F. Surlyk & J. Ineson (eds.): The Jurassic of Denmark, Greenland and adjacent areas, in press. Geology of Denmark Survey Bulletin
- Lidmar-Bergström, K., Olsson, S., & Olvmo, M. (1997). Palaeosurfaces and associated saprolites in southern Sweden. Geological Society, London, Special Publications, 120(1), 95–124.
- Ringberg, B. (1984). Cyclic lamination in proximal varves reflecting the length of summers during late Weichsel in southernmost Sweden. In Climatic Changes on a Yearly to Millennial Basis (pp. 57-62). Springer, Dordrecht.
- Bomfleur, B., McLoughlin, S., & Vajda, V. (2014). Fossilized nuclei and chromosomes reveal 180 million years of genomic stasis in royal ferns. Science, 343(6177), 1376-1377.
- Bomfleur, B., Grimm, G. W., & McLoughlin, S. (2014). A fossil Osmunda from the Jurassic of Sweden—reconciling molecular and fossil evidence in the phylogeny of Osmundaceae. bioRxiv, 005777.
- Vajda, V., McLoughlin, S., & Bomfleur, B. (2014). Fossilfyndet i Korsaröd. In Geologiskt Forum (Vol. 82, pp. 24-29). Geological Society of Sweden.
- Bergelin, I. (2009). Jurassic volcanism in Skåne, southern Sweden, and its relation to coeval regional and global events. GFF, 131(1-2), 165-175.
- Bergelin, I., Obst, K., Söderlund, U., Larsson, K., & Johansson, L. (2011). Mesozoic rift magmatism in the North Sea region: 40 Ar/39 Ar geochronology of Scanian basalts and geochemical constraints. International Journal of Earth Sciences, 100(4), 787-804.
- Tappe, S. (2004). Mesozoic mafic alkaline magmatism of southern Scandinavia. Contributions to Mineralogy and Petrology, 148(3), 312-334.
- Printzlau I, Larsen O (1972) K/Ar determination on alkaline olivine basalts from Skåne, southern Sweden. GFF 94:259–269
- Klingspor I (1973) A preliminary report on the dating by the K/Ar method of the basalt neck at Göbnehall, southern Sweden. GFF 95:287–289
- Klingspor I (1976) Radiometric age-determination of basalts, dolerites and related syenite in Skåne, southern Sweden. GFF 98:195–216
- Bergelin, I. (2006). 40Ar/39Ar Geochronology of Basalts in Scania, S Sweden: Evidence for Two Pulses at 191-178 Ma and 110 Ma, and Their Relation to the Break-up of Pangea. Geologiska institutionen, Lunds univ..
- Tappe, S., Smart, K. A., Stracke, A., Romer, R. L., Prelević, D., & van den Bogaard, P. (2016). Melt evolution beneath a rifted craton edge: 40Ar/39Ar geochronology and Sr–Nd–Hf–Pb isotope systematics of primitive alkaline basalts and lamprophyres from the SW Baltic Shield. Geochimica et Cosmochimica Acta, 173, 1-36.
- Mehlqvist et al., 2009 K. Mehlqvist, V. Vajda, L. Larsson A Jurassic (Pliensbachian) flora from Bornholm, Denmark — a study of a historic plant fossil collection at Lund University, Sweden GFF, 131 (2009), pp. 137-146
- Frandsen, Nils, and Finn Surlyk. 2003. The Lower–Middle Jurassic of the Anholt borehole: implications for the geological evolution of the eastern margin of the Danish Basin. Geological Survey of Denmark and Greenland Bulletin 1. 585–609. Accessed 2020-07-09.
- Erlström, M.; L.O. Boldreel; S. Lindström; L. Kristensen; A. Mathiesen; M.S. Andersen; E. Kamla, and L.H. Nielsen. 2018. Stratigraphy and geothermal assessment of Mesozoic sandstone reservoirs in the Øresund Basin – exemplified by well data and seismic profiles. Bulletin of the Geological Society of Denmark 66. 123–149. Accessed 2020-07-09. ISSN 2245-7070
- McLoughlin, S., & Bomfleur, B. (2016). Biotic interactions in an exceptionally well preserved osmundaceous fern rhizome from the Early Jurassic of Sweden. Palaeogeography, Palaeoclimatology, Palaeoecology, 464, 86-96.
- Tekleva, M. V., Krassilov, V. A., Kvacek, J., & Van Konijnenburg-van Cittert, J. H. A. (2006). Pollen genus Eucommiidites: ultrastructure and affinities. ACTA PALAEOBOTANICA-KRAKOW-, 46(2), 137.
- Crane, P. R. (1996). The fossil history of the Gnetales. International Journal of Plant Sciences, 157(S6), S50-S57.
- Rydin, C., Pedersen, K. R., Crane, P. R., & Friis, E. M. (2006). Former diversity of Ephedra (Gnetales): evidence from early Cretaceous seeds from Portugal and North America. Annals of Botany, 98(1), 123-140.
- Wade-Murphy, J., Kuerschner, W. M., (2006). A new technique to infer the botanical affinity of palynomorphs, and its application on spheripollenites psilatus from the toarcian of bornholm, denmark. In 7 th European Palaeobotany Palynology Conference, Prague (p. 153).
- Bomfleur, B., Grimm, G. W., & McLoughlin, S. (2015). Osmunda pulchella sp. nov. from the Jurassic of Sweden—reconciling molecular and fossil evidence in the phylogeny of modern royal ferns (Osmundaceae). BMC Evolutionary Biology, 15(1), 1-25.
- McLoughlin, S., Bomfleur, B., & Vajda, V. (2014). A phenomenal fossil fern, forgotten for forty years. Deposits Magazine, 40, 16-21.
- Bomfleur, B., Grimm, G. W., & McLoughlin, S. (2017). The fossil Osmundales (Royal Ferns)—a phylogenetic network analysis, revised taxonomy, and evolutionary classification of anatomically preserved trunks and rhizomes. PeerJ, 5, e3433.
- Qu, Y., McLoughlin, N., van Zuilen, M. A., Whitehouse, M., Engdahl, A., & Vajda, V. (2019). Evidence for molecular structural variations in the cytoarchitectures of a Jurassic plant. Geology, 47(4), 325–329. doi:10.1130/g45725.1
| Ph |
| ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|