Sorthat Formation
The Sorthat Formation is a geologic formation on the island on Bornholm, Denmark. It is of latest Pliensbachian-Lower Toarcian age. Plant fossils have been recovered from the formation, along with several traces of invertebrate animals. The Sorthat Formation is overlain by fluvial to lacustrine gravels, along with sands and clay along with parts covered by coal beds, that are part of the Aalenian-Bathonian Bagå Formation.[2] In fact the Sorthat Formation was included on most of late SXX studies as the lowermost part of the Bagå Formation, recovering the latest Pliensbachian to lower Aalenian boundary, that was dismissed after comparing the geology of the two.[3][4] The Sorthat strata recover a mostly marginal deltaic to marine unit, where large streams fluctuated to the east where a large river system was established at the start of the Toarcian.[2] At the northwest the local Vulcanism, that started on the lower Pliensbachian, extended along the North Sea and mostly from southern Sweden.[5] At this time, the Central Skåne Volcanic Province and the Egersund Basin expulsed most of his strata, with influences on the local tectonics.[5] The Egersund Basin has abundant fresh prophyritic Nephelinite lavas and dykes of lower Jurassic Age, with a composition nearly equal to those found on the clay pits. That reveals the translation of strata from the Continental margin by large fluvial channels, that ended on the sea deposits of the Ciechocinek Formation Green series.[5]
| Sorthat Formation Stratigraphic range: Latest Pliensbachian to Toarcian ~183–176 Ma | |
|---|---|
 Korsodde Section of the Sorthat Formation, where the local Toarcian Anoxic event strata is located | |
| Type | Geological formation |
| Underlies | Bagå Formation |
| Overlies | Rønne & Hasle Formations |
| Thickness | 3–200 m (9.8–656.2 ft)[1] |
| Lithology | |
| Primary | Claystone, sandstone[1] |
| Location | |
| Region | Bornholm |
| Country | |
| Type section | |
| Named for | Sorthat-Muleby, Bornholm |
| Named by | Gry (as part of the Bagå Formation) [2] |
| Year defined | 1969 |
 Sorthat Formation (Denmark) | |
Stratigraphy
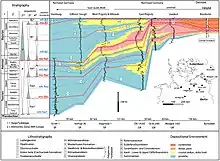
On Bornholm, the lower-middle Jurassic succession is composed of the Rønne (Hettangian-Sinemurian), Hasle (Lower-late Pliensbachian), Sorthat and Bagå Formations. The major Pliensbachian–Bathonian coal-bearing Clays and Sands that overlie the Lower Pliensbachian Hasle Formation are distributed between both the Sorthat Formation and the overlaying Bagå Formation.[1] The Sorthat Formation is the regional equivalent of the Ciechocinek Formation on Baltic Germany and Poland, the Fjerritslev Formation on the Danish Basin, the Rya Formation on Scania.[1] The Sorthat Formation beds where referred originally to the Levka Beds, Sorthat and Bagå beds.[2] The major section of the Formation is the Korsodde coastal section located on the south-west part of the Island.[2] A detailed stratigraphic interpretation of the beds has been difficult to achieve due to the complicated block faulting, but especially due to the absence of marine fossils and distinct marker beds.[2] Using the Megaspore contents, the rocks were dated originally as Middle Jurassic, with the Levka and Sorthat beds being roughly contemporaneous, and the Bagå beds being possibly slightly younger. Latter the Bagå Formation was limited, including the Coal Clays of the Levka Beds, along with the also Coal Dominant beds of the Korsodde and Onsbæk sections.[3] With more advanced Palynological studies recovered from locations such as the Levka-1 core-well and the Korsodde section Upper Pliensbachian strata appeared.[6][7] As at the time several Megaspores collected were found to be common on both the Bagå Formation and Sorthat beds, implying at least the presence of Toarcian-Aalenian strata.[3] Although it was difficult to set a concrete age for the Megaspore-bearing strata.[8] With both, the palynological–sedimentological study of all available exposures and cores from the Lower–Middle Jurassic lead to the revelation that the Hasle Formation (Lower-Middle Pliensbachian) are covered by a succession referable to both the Levka and Sorthat Beds, that are composed mostly by bioturbated Sands, Heteroliths and Clays along with abundant Coal seams containing relatively diverse brackish-marine dinoflagellate assemblages that are indicative of the upper Pliensbachian, Toarcian and possibly lower Aalenian strata.[6] While the upper strata is covered by the fluvial gravels and sands, along with lacustrine clays, carbonaceous clays and coals belonging to the Bagå Formation.[1]
Lithology

The Sorthat Formation has a highly variable lithology.[1] The main core studied from the rocks, the Levka-1 well reveal first a sharp-based, fining-upwards units, 3–14 m thick, consisting of coarse-grained, occasionally pebbly Sand, overlain by muddy, with Coal and Mica, fine- to medium-grained sand, that is laminated to homogeneous Clay and coal seams with roots.[1] On most of the strata there is a common Parallel Lamination with subordinate Cross-bedding, Cross-lamination and Flaser lamination.[1] There are abundant Large plant fragments and small Quartz. Related to this level, there is a clear absence of Marine palynomorphs are not found, as a main probe that this level was deposited on a coastal or Delta plain with fluvial channels, Lakes and Swamps.[6] This is also related to the most recent finds on the German realm of the Ciechocinek Formation, where a large Deltaic System ended. The large Toarcian–Bajocian deltaic systems locally where the shoreline influenced by the vicinity between brackish to freshwater and continental biofacies.[9][10] The North German Basin shows that on approximately 14.4 m.a, four third-order relative sea-level fluctuations led the subsequent formation of four individual delta generations in the Bifrons-Thouarsense (Toarcian), Murchisonae-Bradfordensis (Aalenian) and Humpresianum-Garatiana (Bajocian).[9] The Toarcian section was dominated by regressive elongated river-dominated deltas, where due to the fall of the sea level the south to southwest directed delta progradation between the Lower-Upper Toarcian, that was deposited as 40 m of deltaic successions, found on places like Prignitz (East) and Brandenburg (North).[9] Most of the Palynomorphs found on the Toarcian strata are connected with the ones found on the Sorthat Formation.[9] With nearly 40 m thick, the upper section of the formation is composed mostly by a series of cross-bedded, cross-laminated, wave-rippled and bioturbated sand and heteroliths with sporadic Syneresis cracks, Pyrite nodules, the ichnofossils Planolites isp. and Teichichnus isp. and brackish-marine palynomorphs, mostly dinoflagellates.[1] This upper part has a strata more common to be deposited on nearshore environments with abundant lagoons, coastal lakes and fluvial channels with the clean sand at the top probably representing a marine shoreface.[1] The Korsodde Section, with 93 m thick is composed mostly by coarse-grained sands with cross-bedding and parallel lamination, being mostly of Black due to an abundant organic debris.[1] This Section has been interpreted as part of the large local fluvial system, probably as a series of minor fluvial channels, that were connected with coastal lakes and lagoons, where riparian vegetation was relatively abundant, judging by the presence of megaflora remains and palynomorphs.[1] Small ichnofossil burrows and larger burrows, including Diplocraterion isp. are common, interpreting that there was at least one subunit that was the fill of an estuarine channel.[1] The uppermost part of the formation in the Korsodde section consists of fine-grained, sands with cross stratification and parallel-lamination, yellowish–brown color, along with Sandstones with thin bioturbated and wave-rippled heterolithic beds.[1]
Profile
At Korsodde, the Evironment Includes:
| Unit | Lithology | Thickness (metres) | Type of Environment | Fossil Flora | Fossil Fauna |
|---|---|---|---|---|---|
|
Unit A |
Composed of yellow, weakly cemented Muscovite Quartz sandstone, medium/fine-grained in the lower part, fine-grained in the upper part. Ripple/herringbone lamination is present in most of the beds, along discontinuous mudstone drapes around (0.5 cm thick) and mudstone intraclasts. The mustones show often ferruginization. A single thin horizon which occurs at about 85 cm of the section and also a thin erosional surface with mudstones occurs at 1 m. There is a layer of heterolithic deposits, with fine grained ripple Mudstones and sandstones at 1.65–1.75 m. |
0.45–2.3 m |
Estuarine channel fill (upper or marginal, less energetic part) |
Non Recovered |
|
|
Unit B |
A layer made of intercalations of muscovite quartz sandstones and dark mudstone drapes, with abundant heteroliths. In the vertical section, the sandstone layers (3 cm thick) are lenticular, with some displaying ripple cross and herringbone lamination, and the mudstone drapes (0,5 cm thick) have wavy lamination. This last ones have a few laminae separated by thicker, coarser, mainly silty laminae, showing abundant ferruginous cementation. There is a layer over B considered transitional to C. |
2.3–3.41 m |
Upper tidal flat deposits surrounding an estuary |
Non Recovered |
|
|
Unit C |
It has Two Major layers: a series of 20 cm dark mudstone with horizontal lamination and silt intercalations and a series of dark heteroliths, with intercalated mudstones and ripple limestones. |
3.41–3.7 m |
Restricted bay passing into upper tidal flat deposits |
Non Recovered |
|
|
Unit D |
Yellow ripple cross sandstone with abundant muscovite, alternating with continuous and discontinuous dark mudstone with abundant organic material. There are Pyrite concretions in the lower part. |
3.7–4.7 m |
Lower tidal flat within an estuary |
Roots |
|
|
Unit E |
Composed mostly by fine grained sediments with abundant organic matter. Starts with 55 cm of muddy sandstone, dark at the beginning and ligth in the upper part. A bed of 5 cm of Mudstone overlays the sandstone, followed by various levels of fine-grained sandstones interbedded with dark siltstone-mudstone, pyrite concretions and sandy mudstone. Over this is developed a massive coal layer containing Neocalamites stems where pyrite become more common. It is overlayed by mudstone, fine sandstone that turn into a poorly sorted yellow ferruginous layer. The upper part, with 85 cm thick is composed by mudstone, with allochthonous Neocalamites stems and lignite clasts. |
4.7–6.9 m |
Lagoonal environment above a coal bed |
|
|
|
Unit F |
Mostly pale, fine-grained, ripple cross muddy sandstone and normal sandstone, separated by thin pale sandy mudstones or thin mudstone drapes. Pyrite concretions and lignite clasts occur in the sandstones. There are Synaeresis cracks, noted at 8.15-8.75 m. |
6.9–9.9 m |
Tidal flat deposits in an estuary. |
|
|
|
Unit G |
It starts with a prominent erosional surface, composed by yellow medium/fine-grained cross-laminated sandstones with moscovite. |
9.9–11.35 m |
Estuarine bar |
Non Reported |
Non Reported |
|
Unit H |
Pale fine-grained ripple and herringbone sandstones and mudstones, with intercalations of sandy mudstones and mudstone drapes with intense ferruginization, with some layers of mudstone-sandstone heteroliths |
11.35–14.2 m |
Marginal part of an estuary channel fill |
Non Reported |
|
|
Unit I-J |
Totally bioturbated muddy sandstone |
14.2–14.4 m |
Short-lived bay or lagoon |
|
|
Palynostratigraphy
One of the most complete floras found in Europe dating to the Pliensbachian-Toarcian boundary, and among all the Jurassic palynologycal deposits found worldwide.[4][7][8][12]
Environment
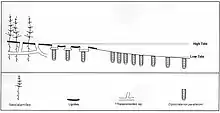
Due to a Late Pliensbachian marine regression, deposition of coal-bearing strata on the Sorthat Formation was resumed on Bornholm until an Early Toarcian transgression terminated peat formation.[13] The two main deposits of the formation, the Levka-1 Well and the lower part of the Korsodde Section where deposited on an environment influenced by the sea, being the Levka location populated by Lagoons, lakes, channels and low fluvial areas.[13] The Bifrons to Levesquei Zone in the coeval units at the east and west of Prignitz a sandy coastal-deltaic succession was replaced by laminated shales with pelagic marine fauna, reflected on the shoreline shifts on the Northeast, what contributed to retrogradational stratal pattern architectures.[14] On the Sorthat Formation is present a transition from upper to lower shoreface environments, indicating a deepening trend. In the Younger Levesquei subzone delta plain environments were replaced by shoreface setting with active bioturbation and hummocky cross-stratification.[14] The Rǿnne Graben shows seismic lines with onlapping patterns that have been correlated to this Lower Toarcian marine shoreface deposits with intense bioturbation.[15]
The depositional environments Include:
- The Levka beds start overliying the foreshore deposits of the Hasle Formation.[16] Its deposition is composed mostly by interbedded sand, clay and coal beds. The loose sand constitute the major parts of the unrecovered intervals.[17] This sand is fine/medium-grained, micaceous, very carbonaceous and muddy, showing mostly parallel lamination, with rare cross-bedding, cross-lamination and flaser lamination.[17] This first levels are interpreted as fluvial channel fills, reflecting active channel deposition followed by decreasing current strength and abandonment with a passive phase of clay deposition, final overgrowth and change into peat-forming swamps.[17] Between the channel fills are intervals with thinly interbedded sand and clay and very common occurrence of rootlets, coal seams and the rapid facies changes, interpreted as representing wet, vegetated, floodplain areas with shallow lakes, swamps and small crevasse deltas receiving overbank spills from nearby active channels.[17] The coal seams analisis revelated that the peatforming swamps were water-saturated, densely vegetated, anoxic and nutrient-rich.[17] It was followed by a coastal or lower delta plain environment, populated by abundant large fluvial channels or distributaries, and nearby floodplain areas where lacustrine-lagoonal mud, crevasse splays and peat accumulated.[17] Later a rise in the sea level is reported due to the increase of Acritarchs and Tasmanites, where a lagoon succession overlain by the fill of a coastal lake that developed into a paleosol.[17] Later, marine palynomorphs become absent and the location become again a crevasse delta and fill of an abandoned fluvial channel, intercalated with lake deposits.[16] After this, a lagoon succession is marked with the appearance of Planolites and Teichichnus burrows and dinoflagellate cysts Nannoceratopsis gracilis, N. senex and N. triceras, and common Botryococcus, indicating a major marine rise event.[16]
- Sorthat Beds where on all its deposition there was a series of intercalated minor or major extended lower delta plain environment deposits, with pyrite nodules and the trace fossil Arenicolites.[16]
- Baga Beds are filled by interbedded sand, clay, and coal beds with rootlets, along plant fragments including small logs, stems and leaves.[16] It represents gradual infill of lakes which were transformed into vegetated swamps where peat accumulated, with isolated sand beds that where deposited in shallow lakes.[17] The increase of roots, suggest he transition to crevasse deltas formed by the amalgamation of crevasse splay deposits and infill of crevasse channels.[16] Some sand beds originated probably on the granite horst located a few hundred meters east of the exposure.[2] There is also outwash material transported over short distances from nearby source areas and rapidly deposited in ponds or lakes.[17] Younger lacustrine clay suggest re-establishment of the lakes, with pyrite as evidence of increased marine influx.[17]
- Korsodde Section overlies the fine-grained sandstones of the Hasle Formation, deposited on a high energy shoreface environment.[17] This section of the formation started as a derived of sand units deposited in fluvial channels, with abundant carbonaceous matter was probably derived from extensive erosion of peat accumulations during changes in channel courses, as proven by the abundant presence of rootlet and coaly beds.[17] The intrusion of younger clay beds led to a gradual infilling of relatively small coastal lakes and enclosed lagoons which became vegetated and turned into peat-accumulating environments (isolated from active clastic sedimentation), eventually forming palaeosols. This units are filled with pyrite nodules and medium-large wood fragments, while the genera Botryococcus, Lecaniella and Mendicodinium reticulatum occur in varying amounts ranging from abundant to rare, and a few acritarchs are also present.[16] Is overlied by the intercalation of crevasse delta deposits and lacustrine-brackish flooding surfaces with shifts between ordinate and subordinate tidal currents, with scattered small burrows (Diplocraterion) and mud drapes on foresets containing abundant Nannoceratopsis senex.[16] Mancodinium semitabulatum and Mendicodinium groenlandicum are also found in this sections, but subordinated to the inner fluvial dominated part of an estuarine channel, overlied by a tidal dominated part.[16] Lagoons of various conditions on younger deposits suggest sea level rise, intercalated with riverine deposits, on a series of regression-transgression trends.[16]
Inertinite has been recovered from the Coal-Bearing levels of the formation, where the palynology shows that the mire vegetation may have been dominated by gymnospermous plants and a secondary proportion of ferns characterised by the genera Dicksonia or Coniopteris and the family Osmundaceae.[13] O several coal seams there where found several Biomolecules, where Euulminite and Attrinite were the most abundant huminite macerals recovered.[18] The Levka-1 well section represents fluvial channels, floodplain areas with shallow lakes and lagoons, and small crevasse deltas, with abundant coalified wood fragments and stems, being most of them found associated to sandy channel fills and on heavily rooted crevasse and lake deposits in shallow inter-fluvial areas.[19] On the Toarcian at Bornholm strata indicates a warm, humid climate suggested by the large number of plant species from interconnected Jameson Land, and thin cutinised leaves of Podozamites and Equisetales comparable in size to modern subtropical bamboos are thought to reflect favourable conditions for plant growth.[19] There is abundant Coal, which indicates that wildfires occurred in the bog.[19] Wood particles from this section, both charcoalified and unburned (coalified), with a rounded and worn nature on many particles, that implies the influence of greater transportation energies.[20] The Toarcian Erratics from Northern Germany are assigned to Southern Sweden and also to the Sorthat Formation, being suggested as the source area of the Ahrensburg erratics.[21] On the Toarcian, the facies on Sweden the sketchy facies information suggests a general lateral coarsening of the lithofacies on Helsingborg (Rya Formation) and the Sorthat Formation, where there is the probe of the presence of a large deltaic system that fluctuated from the SE of Sweden and Bornholm, that ended on a prodeltaic facies belt stretching from Vorpommern to Mecklenburg Bay.[21]
Coal

The Levka- 1 well has a core of approx. 150 m on the Sorthat Formation, covering the underlying marine strata of the Hasle Formation.[22] The lower part includes 112 m with the Coal along sand and clay. There if an abundant presence of large, coalified, wood fragments and stems.[22] The Coal bearing strata of the Levka-1 are interpreted as fluvial channel fills, with active channel deposition followed by abandonment and a passive phase of clay deposition, gradual overgrowth and change into a peat-forming environment.[22] Clay and coal seams present along this strata have abundant rootlets and a non-marine palynomorph assemblage dominated by spores and pollen, interpreted as representing flood plain areas with shallow Lakes, small crevasse Deltas and Swamps. Some sections have wave ripples, wavy and flaser bedding, bioturbation and transported Equisetites stems, that are interpreted as the sediment fill of a local lagoon, deposited on a transgressive shoreline with a series of lagoon successions.[22] Levka-1 coal contain hard, black coal and are very similar petrographically, with Huminite on the major part of the seams, with some seams having up to 90% of Huminite. There is a dominance of macerals from detrital organic matter (Humodetrinite) over macerals derived from more woody material (Humotelinite).[22] Gelinite appears as the most common component of the samples, followed by Huminite.[22]
The Korsodde section is interpreted as representing a small coastal series of lakes and protected lagoons, where are recovered at least 6 coal seams occur. Above the marine strata of the Toarcian Tranggression strata with abundant Clay fine-grained Sand, Silt, that contains transported, coalified pieces of wood, pyrite nodules, rootlets and a diverse microspore assemblage, where there is an abundance of the marine dinoflagellate Mendicodinium reticulaturn.[22] On this coal Seams Huminite maceral group comprises the majority of the organic matter, where humotelinite dominates over humodetrinite maceral. Eu-ulminite and Densinite are the most prominent macerals.[22]
Fungi
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Fungus Spores of Incertain Classification. The three uppermost samples of the Korssode Section are poor in diversity, with Fungal spores are common in at least one sample; these have not been recorded from the samples below. |
||
Phytoplankton
On the Lower Jurassic of Bornholm there where several successions of nearshore Peat Formations with Dinoflajellates.[22] Coal-bearing strata were deposited in an overall coastal plain environment during the Hettangian-Sinemurian and then during the Early Pliensbachian was interrupted until the late Pliensbachian-Lowermost Toarcian due to a Sea regression.[22]
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Type Genus of the Botryococcaceae inside Trebouxiales. A colonial green microalga related with freshwater and brackish ponds and lakes around the world, where it often can be found in large floating masses. Sorthat Formation Botryococcus lived on an environment interpreted as a coastal lake, permanently vegetated and shallow that was occasionally flooded by the sea. |
 Specimen | |
|
|
|
Affinities with the family Zygnemataceae. A Genus derived from Freshwater filamentous or unicellular, uniseriate (unbranched) green algae. |
||
|
Ovoidites[16] |
|
|
|
Affinities with the family Zygnemataceae. A Genus derived from Freshwater filamentous or unicellular, uniseriate (unbranched) green algae. |
|
|
Crassosphaera[16] |
|
|
|
Affinities with the family Pycnococcaceae. |
|
|
Pterospermella[16] |
|
|
|
Affinities with the family Pterospermataceae. |
|
|
Pterospermella[16] |
|
|
|
Affinities with the family Pterospermataceae. |
|
|
Leiosphaerida[16] |
|
|
|
Affinities with the family Prasinophyceae. |
|
|
Cymatiosphaera[16] |
|
|
|
Affinities with the family Pterospermopsidaceae. |
|
|
Tasmanites[16] |
|
|
|
Affinities with the family Pyramimonadaceae. Found on shoreface and shoreface-offshore transition zone deposits. |
|
|
Veryhachium[16] |
|
|
|
A Dinoflajellate, member of the Dinophyceae. |
|
|
Dissilodinium[16] |
|
|
|
A Dinoflajellate, member of the Gymnodiniales. |
|
|
Luehndea[16] |
|
|
|
A Dinoflajellate, member of the Luehndeoideae. It stablishes the Luehndea spinosa Zone; the age of this zone is late Pliensbachian to early Toarcian. |
|
|
Korystocysta[16] |
|
|
|
A Dinoflajellate, member of the Cribroperidinioideae. |
|
|
|
|
A Dinoflajellate, type Genus of the Mancodinioideae. |
||
|
|
|
A Dinoflajellate, member of the family Gonyaulacales. |
||
|
|
|
A Dinoflajellate, member of the family Nannoceratopsiaceae. It is a genus related with Marine deposits. The presence of Nannoceratopsis gracilis, N. senex and N. triceras, and common Botryococcus is interpreted as a lagoon succession overlying a transgressive surface and indicates a rise in relative sea level. |
||
|
Micrhystridium[16] |
|
|
|
An Acritarch, familia incertae sedis |
|
Terrestrial palynology
In Early Toarcian carbonates local bulk organic matter, and wood fragments have been related to carbon cycle perturbations, helping to know the reaction of the Continental Biota to the TAOE that happened along contemporaneous large scale volcanism.[24] There where several changes on the woody vegetation in the wood-derived carbon, with pollen assemblages are dominated by Taxodiaceae pollen types such Cerebropollenites associated with Cycad pollen types (Chasmatosporites) and the Cheirolepidiaceous Corollina, reflecting a warming transition from a wet fern dominated landscape to seasonally dry vegetation.[24] Is also common to found wood from a near-shore setting on the eastern Danish Basin, with two sets: macroscopic wood, recognizable to the naked eye, up to branch-sized; and microscopic wood (0.25 to 1 mm average dimension).[25] Apart from wood and leaf cuticles there is little organic matter present in the section—the sediments comprise sand, silt and mud—and thus there is no possibility of contamination by marine carbon.[25]
Bryophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Incertade Sedis affinities with Bryophyta. This Spore is found on the Jurassic Sediments associated with the Polar Regions. The Sorthat Formation is among its southernmost locations. |
||
|
|
|
Incertade Sedis affinities with the Bryophyta. Uncertain affinity possible Moss Spores. This Genus can Also be derived from a Lycophyte. |
||
|
|
|
Incertade Sedis affinities with the Bryophyta. Uncertain affinity possible Moss Spores. This Genus is Know From Paleozoix Strata, so it can be a reworked genus from older Layers. |
||
|
|
|
Affinities with the family Notothyladaceae inside Anthocerotopsida. Hornwort spores. |
).png.webp) Example of extant Notothylas specimens, Foraminisporis come probably from similar genera | |
|
|
| |||
|
|
| |||
|
|
| |||
|
|
|
Affinities with the family Sphagnaceae inside Sphagnopsida. "Peat moss" spores, relted to genera such as Sphagnum that can store water large amount of water. |
 Example of extant Sphagnum specimens, Stereisporites, Sculptisporis and Rogalskaisporites come probably from similar genera | |
|
|
| |||
|
|
| |||
|
|
|
Affinities with the family Encalyptaceae inside Bryopsida. Branching Moss Spores, related with high water-depleting environments |
_2281.JPG.webp) Example of extant Encalypta specimens, Staplinisporites come probably from similar genera | |
|
|
|
Affinities with the Sphaerocarpales inside Marchantiopsida. Liverwort spores, related to high humid environments |
 Example of extant Sphaerocarpos specimens, Aequitriradites come probably from similar genera | |
Lycophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with the family Lycopodiaceae inside Lycopodiopsida. Lycopod spores, related with herbaceous to arbustive flora common on humid environments |
 155pxExtant Lycopodium specimens. Genera like Sestrosporites, Camarozonosporites, Retitriletes, Lycopodiumsporites and Semiretisporis probably come from a similar plant | |
|
|
| |||
|
|
| |||
|
Lycopodiumsporites[26] |
|
|
| ||
|
|
| |||
|
|
|
Affinities with the families Sigillariaceae and Lepidodendraceae inside Lycopodiopsida. Basal Pseudofern Arboreal Plant Spores, probably Relict Genus or Reworked Spores from Older Layers |
||
|
|
| |||
|
|
|
Affinities with the Isoetales inside Lycophyta. Low floor spores, related with herbaceous to arbustive flora common on humid environments |
 Extant Isoetes specimens. Genera like Minerisporites, Echitriletes and Paxillitriletes probably come from a similar plant | |
|
|
| |||
|
|
| |||
|
|
|
Affinities with Pleuromeiales inside Lycophyta. The Plueromeiales were tall Lycophites (2 to 6 m) common on the Trassic. Probably come from a relict genus. |
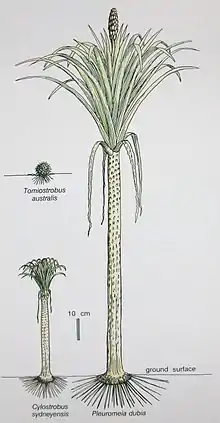 Reconstruction of the extinct genus Pleuromeia, typical example of Pleuromeiales. Nathorstisporites and Aratrisporites probably come from a similar or a related Plant | |
|
|
| |||
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. Herbaceous Lycophyte flora, similar to Ferns, ralated with Humid Settings. This Family of Spores are also the most diverse on the Formation. |
 Extant Selaginella, typical example of Selaginellaceae. Genera like Anapiculatisporites, Limbosporites, Cabochonicus, Kraeuselisporites, Hughesisporites, Erlansonisporites,Apiculatisporis, Bacutriletes,Perotrilites,Horstisporites, Maexisporites, Aneuletes, Neoraistrickia, Trileites, Verrutriletes, Cadargasporites and Densoisporites probably come from a similar or a related Plant | |
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
Sphenophyllaceae
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with the family Cheirostrobaceae inside Sphenophyllales. It was tougth to be related to the basal plants of the Devonian, such as the genus Rhynia. Later, representatives from the genus where recovered with the sporangiophores of the cone genus Cheirostrobus. Probably represent Miospores from a relict Genus. |
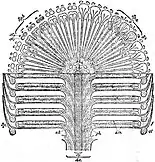 Diagram of the Cone Cheirostrobus, found associated with Retusotriletes miospores | |
Equisetales
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with the Calamitaceae inside Equisetales. Horsetails, herbaceous flora related to high humid environments, flooding tolerant plants. On the sections of the Formation such as Korsodde this genus has small peaks in abundance at the layers where more Equisetites stems are found. |
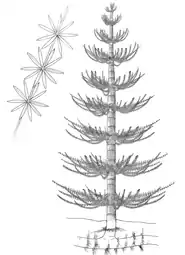 Recosntruction of the Genus Calamites, found associated with Calamospora | |
Gymnospermophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Incertade sedis Affinities with the Gymnospermophyta. Spores of Uncertain provenance, that can be derived from any of the members of the Gymnospermophyta. The lack o distinctive characters and bad conservation are among the main factors to make this Palynological residues difficult to classify. |
||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
|
Pollen from the Family Caytoniaceae inside Caytoniales |
||
|
|
| |||
Pteridophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Incertade Sedis affinities with the Pteridophyta. Uncertain Pteridophyte origin |
||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
|
Affinities with the Callistophytaceae inside Callistophytales. Spores from large arboreal to arbustive ferns |
||
|
|
|
Affinities with the family Botryopteridaceae inside Polypodiopsida. It comprises the main megaspore zonation of the Toarcian of Denmark, being the most abundant spore found. |
||
|
|
|
Affinities with the Ophioglossaceae inside Filicales. Fern spores from lower herbaceous flora |
 Example of extant Helminthostachys specimens, Lycopodiacidites come probably from similar genera or maybe an species from the genus | |
|
|
|
Affinities with the family Lygodiaceae inside Polypodiopsida. Climbing fern spores |
 Example of extant Lygodium, Lygodioisporites come probably from similar genera or maybe an species from the genus | |
|
|
| |||
|
|
|
Affinities with the family Dennstaedtiaceae inside Polypodiales. Forest Fern Spores |
.jpg.webp) Example of extant Dennstaedtia specimens, Leptolepidites come probably from similar genera | |
|
Osmundacidites[26] |
|
|
|
Affinities with the family Osmundaceae inside Polypodiopsida. Near Fluvial currents ferns, reted to the modern Osmunda Regalis. |
 Example of extant Osmunda specimens, Osmundacidites and Baculatisporites come probably from similar genera or maybe an species from the genus |
|
|
| |||
|
|
| |||
|
|
|
Affinities with the Pteridaceae inside Polypodiopsida. Forest Ferns from humid ground locations |
.jpg.webp) Example of extant Pityrogramma specimens, Contignisporites and Manumia come probably from similar genera or maybe an species from the genus | |
|
|
| |||
|
|
|
Affinities with the genus Ceratopteris (Parkerioideae) inside Polypodiopsida. Spores from aquatic or subaquatic ferns of moderate size. Are abundant on the layers near fluvial deposition, probably associated to large underwater colonies of ferns on local freshwater deposits. |
 Example of extant Ceratopteris specimens, Striatriletes come probably from similar genera or maybe an species from the genus | |
|
|
|
Affinities with the Fern Genus Saccoloma, type representative of the family Saccolomataceae. This Fern Spore has resemblance only with the living genus Saccoloma, being probably from a Pantropical genus found in wet, shaded forest areas. |
 Example of extant Saccoloma specimens, Annulispora come probably from similar genera or maybe an species from the genus | |
|
|
|
Affinities with the family Cyatheaceae inside Cyatheales. Arboreal Fern Spores |
 | |
|
|
| |||
|
|
| |||
|
|
|
Affinities with the genus Dicksoniaceae inside Polypodiopsida. Tree fern spores |
_C.Chr._by_Jason_Hollinger_001.jpg.webp) Example of extant Lophosoria specimens, Tripartina and Undulatisporites come probably from similar genera | |
|
Undulatisporites[26] |
|
|
| ||
|
|
|
Affinities with the Gleicheniales inside Polypodiopsida. Fern spores from lower herbaceous flora |
 Example of extant Gleichenia specimens, Gleicheniidites and Iraqispora come probably from similar genera or maybe an species from the genus | |
|
|
| |||
|
|
|
Affinities with the Marattiaceae inside Polypodiopsida. Fern spores from lower herbaceous flora |
.jpg.webp) Example of extant Marattia specimens, Marattisporites come probably from similar genera | |
|
|
|
Affinities with the Matoniaceae inside Polypodiopsida. Fern spores from lower herbaceous flora |
 Example of extant Matonia specimens, Matonisporites come probably from similar genera | |
Gnetales
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Type Pollen of the Erdtmanithecales, that can be related with the Gnetales. Thick tectum, infratectum of small granules, indistinct or absent foot layer. Originally was thought to come from Angiosperms, latter reports suggest it come from arbustive Bennetites.[28] It was recently found to come from Eucommiitheca, member of the enigmatic Erdtmanithecales, reinterpreted as an unusual gymnosperm grain with a single distal colpus flanked by two subsidiary lateral colps.[29] Is very similar to the Pollen of the extant Ephedra and Welwitschia (mainly on the granular structure of the exine).[30] |
||
|
|
|
Affinities with Gnetopsida and probably Gnetophyta. Has Been considered Pollen of Chloranthaceae. However, it is to old for belonging to advanced Angiosperms. It probably comes from cones related to the Genera Piroconites kuesperti from the Lowermost Jurassic of Germany, resembling pollen of extant Ephedra and Welwitschia. |
 Closer Look of Ephedra cones, a common Gnetal. Monosulcites and Clavatipollenites maybe come from a related plant | |
|
|
| |||
Bennettitales
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with the family Cycadeoidaceae inside Bennettitales. Coming From Low Arbustive Bennetites, similar to modern Cycas of the Genus Zamia. This pollen is found related with Dry environments. |
||
Coniferophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with the family Pinaceae inside Pinopsida. Conifer pollen from medium to large arboreal plants |
 | |
|
|
| |||
|
|
| |||
|
|
| |||
|
|
|
Affinities with the cone genus Masculostrobus, a member of the Krassiloviaceae. This Genus has been found associated with Masculostrobus cones on the Middle Jurassic Strata of Purbeck, England. Represents Pollen from Dry Environment-Derived Arboreal-Arbustive Flora. |
||
|
|
|
Affinities with the Podocarpaceae inside Pinopsida. Conifer pollen from medium to large arboreal plants |
.jpg.webp) | |
|
|
| |||
|
|
|
Affinities with the family Cupressaceae inside Pinopsida. Pollen that resembles extant genera such as the Genus Actinostrobus and Austrocedrus, probably derived from Dry environments. |
 | |
|
|
| |||
|
|
|
Affinities with the Cheirolepidiaceae inside Pinopsida. Spheripollenites can compose up to 95% of an assemblage and is correlated with Toarcian carbon cycle anomalies including the oceanic anoxic event.[26] Rhaetipollis is less abundant. Both are genera that indicate extremely dry environments. |
||
|
|
| |||
|
|
| |||
|
|
|
Affinities with the family Araucariaceae inside Pinales. Conifer Pollen from medium to large Arboreal Plants |
 | |
|
|
| |||
Plant macrofossils
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Equisetaceae inside Equisetales. Equisetalean Stems, related that are also found on the Hettangian strata along Skane, Sweden. On the Lagoonar sections there is correlation between bioturbation and transported Equisetites stems.[32] Local Equisetales obtained a considerable size, comparable to modern subtropical bamboos, close to lakes and in the wettest environments.[22] |
 Equisetites specimen | |
|
|
|
Affinities with Calamitaceae inside Equisetales. Equisetalean Stems, related that are also found on same age strata along Skane, Sweden. Based on analogies with morphologically similar extant Equisetum species, it is interpreted to represent a plant of consistently moist habitats, such as marshes, lake margins or forest understorey, developed normally dense thickets. |
 Neocalamites specimen | |
|
|
|
Affinities with Equisetidae inside Equisetales. |
||
|
|
|
Affinities with Lycopodiidae inside Lycopodiales. |
||
|
|
|
Incertae Ordinis inside Pteridophyta. Spiropteris is the name given to the fossil of a fern leaf before opening, when it is still coiled. |
||
|
|
|
Affinities with Osmundaceae inside Osmundales. Related with species commonly reported from the Triassic–Jurassic of southern Sweden. |
 Cladophlebis nebbensis specimen | |
|
|
|
Affinities with Osmundaceae inside Osmundales. |
||
|
|
|
Affinities with Aspleniaceae inside Blechnales. |
||
|
|
|
Affinities with Dicksoniaceae inside Cyatheales. The Lund-material is dominated by ferns belonging to the genus Eboracia (28 specimens of E. lobifolia and 14 of E. sp.). Eboracia sp. has considerably smaller pinnules than lobifolia. |
||
|
|
|
Affinities with Dicksoniaceae inside Cyatheales. Common cosmopolitan Mesozoic Tree fern genus. |
||
|
|
|
Affinities with Dicksoniaceae inside Cyatheales. It show similarities with Sphenopteris longipinnata on the morphological outline of the leaflets and the keels of the pinnate axis. |
||
|
|
|
Affinities with Dipteridaceae inside Polypodiales. Specimens from the same species have been found on the same age Höör Sandstone at southern Sweden. |
||
|
|
|
Affinities with Dipteridaceae inside Polypodiales. |
||
|
|
|
Affinities with Dipteridaceae inside Polypodiales. Dictyophyllum is a common Dipteridacean genus of the mid-Mesozoic |
 Dictyophyllum nilssonii specimen | |
|
|
|
Affinities with Dipteridaceae inside Polypodiales. |
||
|
|
|
Affinities with Matoniaceae inside Gleicheniales. |
||
|
|
|
Affinities with Matoniaceae inside Gleicheniales. |
||
|
|
|
Affinities with Marattiaceae inside Marattiopsida. |
||
|
|
|
Affinities with Caytoniaceae inside Pteridospermatophyta. Related with the Cycadophytes present in the Rhaetic flora of Sweden. |
 Sagenopteris specimen | |
|
|
|
Affinities with Caytoniaceae inside Pteridospermatophyta. Less common that other Arboreal Plants |
||
|
|
|
Affinities with Corystospermaceae inside Pteridospermatophyta. |
||
|
|
|
Affinities with Umkomasiaceae inside Pteridospermatophyta. |
||
|
|
|
Affinities with Williamsoniaceae inside Bennettitales. Insufficient and incomplete material prevents certain allocation to that species. |
.jpg.webp) Otozamites specimen | |
|
|
|
Affinities with Williamsoniaceae inside Bennettitales. |
||
|
|
|
Affinities with Williamsoniaceae inside Bennettitales. |
||
|
|
|
Affinities with Cycadeoidaceae inside Bennettitales. The Most common and abundant Bennetite on the formation. |
.jpg.webp) Nilssonia specimen | |
|
Nilssoniopteris[41] |
|
|
|
Affinities with Cycadeoidaceae inside Bennettitales. |
|
|
|
|
Affinities with Cycadales inside Cycadopsida. |
||
|
|
|
Affinities with Gnetopsida inside Gnetophyta. |
||
|
|
|
Affinities with Ginkgoaceae inside Ginkgoales. Seven species assigned to either Ginkgo or Ginkgoites have been reported from the Latest Triassic to middle Jurassic strata of southern Sweden. |
 Ginkgoites specimen | |
|
|
|
Affinities with Ginkgoaceae inside Ginkgoales. Unlike other Plants specimens from the locations is linked more to Middle Jurassic Flora |
 Baiera specimen | |
|
|
|
Affinities with Czekanowskiales inside Ginkgoales. This Genus is related to flora from the Rhaetian-Hettangian boundary of Jameson Land, but also present on Romania. |
||
|
|
|
Affinities with Czekanowskiales inside Ginkgoales. |
||
|
|
|
Affinities with Czekanowskiales inside Ginkgoales. |
||
|
Solenites[39] |
|
|
|
Affinities with Czekanowskiales inside Ginkgoales. This species was described on the basis of individuals collected in Greenland, from the Triassic-Jurassic boundary. |
|
|
|
|
Affinities with Czekanowskiales inside Ginkgoales. Linked to the Lower Liassic Flora from Greenland. |
||
|
|
|
Affinities with Araucariaceae or Cheirolepidiaceae inside Pinales. P. kurrii (originally P. steenstrupi) is preferred as this species is characterised by relatively broad leaves inserted at high angles to the stem. P. peregrinum has been found on the Hettangian Ronne Formation associated with Cheirolepidaceous wood, Brachyoxylon rotnaensis (Simplicioxylon). On the Toarcian levels, is the most common plant Cuticle recovered locally. |
 Pagiophyllum specimen | |
|
|
|
Affinities with Araucariaceae or Cheirolepidiaceae inside Pinales. Is related to Hettangian axis found on Scania, Sweden |
 Brachyphyllum specimen | |
|
Dactyletrophyllum[31] |
|
|
|
Affinities with Cheirolepidiaceae inside Pinales. Is related to other representatives of the genus of the Toarcian of Italy and Lower Jurassic of Israel. Spheripollenites co-occurs with cuticles of Dactyletrophyllum ramonensis, and after a test of relationships it was found a highly significant correlation may suggest that S. psilatus is produced by the conifer genus Dactyletrophyllum.[31] |
|
|
|
|
Affinities with Cheirolepidiaceae inside Pinales. The main genus of the Cheirolepidaceae, linked with dry environmets and probably Fire Tolerant |
||
|
|
|
Affinities with Palissyaceae inside Pinales. |
||
|
|
|
Affinities with Palissyaceae inside Pinales. Descriptions of the Palissia come mostly from coeval deposits on the Northern Hemisphere, based on very few specimens from Sweden, Germany or America. |
||
|
|
|
|||
|
|
|
Affinities with Krassiloviaceae inside Pinales. The local Podozamites show a rather great range of Growth, reflecting Tropical to subtropical conditions. |
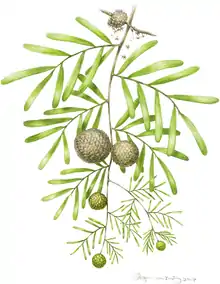 Podozamites reconstruction | |
|
|
|
Affinities with Taxodiaceae inside Pinales. It was originally described as Taxites? subzamioides, later fused with Sewardiodendron. Cunninghamia-like conifers belonging to half-evergreen trees. |
||
|
|
|
Affinities with Taxodiaceae inside Pinales. It matches with the Middle Jurassic Cyparissidium blackii from Yorkshire, England. |
||
Fauna
Ichnofossils
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Dwelling traces |
Marine, Brackish or Freshwater Unbranched U-shaped burrows having a subvertical orientation, with or without lining and passive fill. Are common on modern coastal environments, done by deposit-feeding polychaetes of the families Spionida and Carpitellida, as well as by suspension-feeding Amphipodan crustaceans and deposit-feeding Sipuncula. On this section are recovered only on lower delta plain environment deposits. |
 | |
|
|
Cylindrical burrows |
Burrow-like ichnofossils. It is referred to vermiform deposit-feeders, mainly Polychaetes, producing active Fodinichnia. It is controversial, since is considered a strictly a junior synonym of Palaeophycus.[44] |
 Example of Planolites fossil | |
|
Palaeophycus[11] |
|
|
Cylindrical, predominantly horizontal to inclined burrows |
Burrow-like ichnofossils. They occur in different size classes, 3, 5 and 10 mm in diameter. Are mostly produced by predaceous Polychaeta in marine environments, but other makers may have been involved in continental settings, such as semiaquatic Insects (Orthoptera and Hemiptera) or semiaquatic and non-aquatic Beetles. |
 Example of Palaeophycus fossil |
|
Bornichnus[11] |
|
|
Tubular Traces |
Burrow-like ichnofossils. Exclusive from the Formation, Bornichnus differs from Palaeophycus Hall in its tangled, contorted morphology and abundant branching. Small open burrows produced probably by farming worm-like animals (Probably Polychaeta). Similar complicated burrow systems are produced by the polychaete Capitomastus cf. aciculatus. |
|
|
|
Cylindrical to subcylindrical Burrows |
Burrow-like ichnofossils. Ichnofossils done by organisms advancing along the bottom surface. Very narrow, vertical or subvertical, slightly winding unlined shafts filled with mud. Interpreted as dwelling structures of vermiform animals, more concretely the Domichnion of a suspension-feeding Worm or Phoronidan, with certain Skolithos representing entrance shafts to more complicated burrows. |
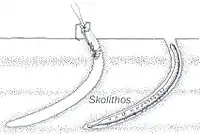 Skolithos ichnofosil reconstruction, with possible fauna associated | |
|
Cylindrichnus[11] |
|
|
Burrowing and track ichnofossils. |
Burrow-like ichnofossils. Cylindrichnus isp. was found only in the uppermost part of the section, and probably represents Polychaeta Burrows.[45] |
|
|
|
Dwelling traces |
Burrow-like ichnofossils. The level where this ichnogenus is more abundant is also composed of abundant fragments of Spreite lamination, derived from the intersection with the Ichnofossil. They are believed to be Fodinichnia, with the organism adopting the habit of retracing the same route through varying heights of the sediment, which would allow it to avoid going over the same area. The assigned origin of this trace are Annelid worms. Aside from the classical interpretation of Polychaeta as producers, many features accord with an interpretation of dwelling Echiurans and Holothurians. |
 Example of Teichichnus fossil | |
|
|
Tubular Fodinichnia |
Burrow-like ichnofossils. Large burrow-systems consisting of smooth-walled, essentially cylindrical components. Can be related to Crustaceans (Anomura, Decapoda), Annelids (Polychaeta, Sipuncula) and Fishes (Dipnoi). Is found associated with Teichichnus. |
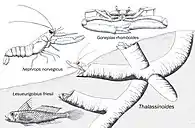 Thalassinoides burrowing structures, with modern related fauna, showing the ecological convergence and the variety of animals that left this Ichnogenus. | |
|
|
Tubular Fodinichnia |
Burrow-like ichnofossils. Interpreted as the feeding burrow of a sediment-ingesting animal.[47] A more recent study has found that Scoloplos armiger and Heteromastus filiformis, occurring in the German Wadden Sea in the lower parts of tidal flats, make burrows that are homonymous with numerous trace fossils of the ichnogenus.[48] |
 Illustration of Chondrites bollensis | |
|
|
Trace Fossil |
Burrow-like ichnofossils. Vertical or oblique complex trace fossil composed of a bunch of spindle-shaped structures and associated tubes, typical of a restricted environment (?estuarine/lagoonal). The local Asterosoma is similar to the Ichnogenus Parahentzschelinia surlyki from the lower Jurassic of Greenland, which can be a junior synonym. This trace fossil is interpreted as made by a small deposit-feeding animal, living in a tube communicated with the sea floor. These traces are linked with shrimps or other aquatic arthropods, since the tunnels possess scratch patterns. |
||
|
|
"U" Shaped Burrows |
Burrow-like ichnofossils. Most Diplocraterion show only protrusive spreit, like the local ones, produced under predominantly erosive conditions where the organism was constantly burrowing deeper into the substrate as sediment was eroded from the top.[49] Most Diplocraterion show only protrusive spreit, like the local ones, produced under predominantly erosive conditions where the organism was constantly burrowing deeper into the substrate as sediment was eroded from the top. "U"-shaped burrows, such as Diplocraterion, can be constructed by a wide variety of creatures: Polychaeta annelids (Axiothella, Abarenicola and Scolecolepis), Sipunculans (Sipunculus), Enteropneustans (Balanoglossus) and Echiurans (Urechis).[50] |
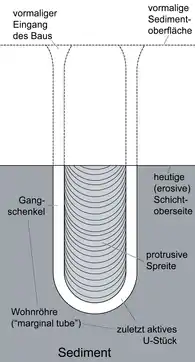 Diplocraterion parallelum diagram | |
Annelida
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
2 Specimens |
A sessile, marine annelid tube worm of the family Serpulidae. Its affinities with the genus Serpula are controversial, since the genus is known mostly since Cretaceous strata. Although there are other fossils assigned to the genus on same age deposits of France.[54] |
 Head of a modern Serpula vermicularis | |
Brachiopoda
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cunchs |
A Saltwater lamp shell Brachiopodan, member of Terebratulidae inside Terebratulida. The specimens are rather incomplete. The genus is a possible junior synonym of Terebratula. |
_flavascens.jpeg.webp) | |
|
|
Cunchs |
A saltwater Brachiopodan, member of Rhynchonellata inside Rhynchonellida. Found associated with Plicatula on long-term well-oxygenated conditions within the substrate and bottom waters. |
| |

Bivalvia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A saltwater Pearl Oyster, member of the family Pteriidae inside Ostreida. Some specimens are big and found associated with Burrow-filled surfaces. This well-known species occurs extremely frequently; Copies occur at all ages, as well as right-handers Left shells have been found. |
||
|
|
|
A saltwater Oyster, member of the family Bakevelliidae inside Ostreida. Found on a series of incomplete Stone Cores filled with specimens, that by Size and Form can be possibly attributable to Gervilleia aerosa. |
| |
|
|
|
A saltwater Clam, type member of the family Parallelodontidae inside Arcida. This species occurs frequently and varies a lot at Korsodde |
||
|
|
|
A saltwater Clam, type member of the family Astartidae inside Carditida. It is found on small, smooth Clam associations, with the exception of one does section, with less well-preserved stone cores and imprints. |
| |
|
|
|
A saltwater Clam, type member of the family Tancrediidae inside Carditida. Tancredia lineata Is based on six shell cores, of which one complete, the others a little defective at the front end. |
||
|
|
|
A saltwater Clam, type member of the family Cardiidae inside Carditida. Is a frequently occurring Clam. |
| |
|
|
|
A saltwater Clam, type member of the family Lucinidae inside Lucinida. Three Stone Cores show perfect Compliance with Regard to Hinge Building and Muscle Impressions Appearance. |
||
|
|
|
A saltwater Clam, type member of the family Pleuromyidae inside Pholadida. Three specimens of this species have been found. |
||
|
|
|
A saltwater file Clam, type member of the family Limidae inside Pectinida. Lima succincta is based on a single fragment of cunch, but enough complete to be certain to be attributed to it. |
| |
|
|
|
A saltwater file Clam, member of the family Limidae inside Pectinida. Mostly know due to a single young individual, but there are also a number of stone cores and imprints of adult Specimens of this species that appear to have been common. |
| |
|
|
|
A saltwater Clam, member of the family Kalenteridae inside Cardiida. One of the most frequent and characteristic Fossils in the layer is this kind, of which especially the hinge part in Imprints and Stone Cores are extremely common. The largest specimen has reached a length of 53 mm, with a width of 39 mm. Myoconcha jespersenii does not occur as frequently as Myoconcha stampensis, with less than 50 specimens, of which, however, none is complete; most occur as stone cores. |
| |
|
|
|
A saltwater nut Clam, type member of the family Nuculidae inside Nuculida. Two copies of a Nucula (leda?) so closely in accordance with Moberg's depictions and description of occurring at Kurremolla. |
| |
|
|
|
A saltwater pointed nut Clam, type member of the family Nuculanidae inside Nuculida. Nuculana bornholmiensis is a very common genus only found locally. Nuculana subovalis occurs less and is highly variable in size and appearance. 10 more or less complete specimens of a large Nuculana species must be attributed to Nuculana complanata. |
| |
|
|
|
A saltwater Scallop, type member of the family Pectinoidae inside Pectinida. This species has been exceedingly frequent, and numerous Fragments are available. Found by some Fragments with the complete cunch preserved have made the provision quite safe. While the presence of Pecten priscus is quite certain, is the Diagnosis of Pecten aequivalvis is a little controversial, due to be based on shell fragments. |
| |
|
|
|
A saltwater Scallop, member of the family Pectinoidae inside Pectinida. A Pair of incomplete specimens of a Hinnites are known and probably match with the species Hinnites tumidus; however, the state of preservation is bad, with the assignment rather uncertain. |
| |
|
|
|
A saltwater Scallop, type member of the family Plicatulidae inside Pectinida. Two poorly preserved specimens have been found. |
||




.jpeg.webp)
.jpeg.webp)


.jpeg.webp)
Scaphopoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A saltwater tusk shell (Scaphopoda), type member of the family Dentaliidae inside Dentaliida. Dentalium etalense is very common on the Transgression deposits of Korsodde. Three specimens are somewhat different from Dentalium etalense, as they have strongly pronounced transverse striation and do not present any curvature. The thickness is also slightly greater than in Dentalium etalense. |
| |

Gastropoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A saltwater Snail, type member of the family Pleurotomariidae inside Pleurotomarioidea. Stone cores are extremely common, now and then imprints of this Art. Of course, it is uncertain, yes often ullmligt, to determine a kind of the stone core alone; but in many cases, the upper side has shown those characteristics of this species. |
| |
|
|
|
A saltwater Snail, type member of the family Turbinidae inside Turbinoidea. Occurs frequently, especially on marine ingression surfaces. |
| |
|
|
|
A saltwater Snail, type member of the family Eucyclidae inside Seguenzioidea. Of this genus there are, in addition to a number of smaller ones, well-preserved stone cores, three imprints. Since the material available is rather poor, is rather impossible to give the specimens found a concrete species assigantion. |
||
|
|
|
A saltwater Snail, member of the family Pyramidellidae inside Pyramidelloidea. Two Specimens of a Turritella-like Gastropod found locally have been refer to the Genus. Chemnitzia citharella includes ten specimens, the largest of the genus found locally. However, most of the specimens are shell cores. |
| |
|
|
|
A saltwater Snail, type member of the family Acteoninidae inside Prosobranchia. Of this small species there are quite a few specimens, both stone cores and imprints that perfectly allow identifying the species. |
||
|
|
|
A saltwater top Snail, type member of the family Trochidae inside Trochoidea. Rare compared with other species found locally. |
||



Crustacea
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A marine/brackish/frashwater Crab, whose classification is problematic, as was suggested to be a member of the family Goniodromitidae, but recent studies suggest affinities with Eubrachyura.[57] Is one of the oldest reported crabs know. Probably lived linked to the parallic deposits of the formation and probably one of the makers of the burrows recovered. It was suggested to come from younger deposits, but recent works confirm a Late Liassic Age.[56] |
||
Chondrichthyes
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A marine/brackish/frashwater Shark, type member of the family Hybodontidae inside Hybodontiformes. Of this genus occurs mostly at Pigstraale, and is in the shape of the cross section is quite similar to Hybodus minor. In addition there is a large amount of teeth, most consistent with the depicted species. Other species occur; between the material where found Teeth that resemble the species Hybodus grossiconus and Hybodus cloacinus |
| |
|
|
|
A marine/brackish/frashwater Shark, type member of the family Acrodontidae inside Hybodontiformes. A large number of Acrodus teeth have been found; they hear fairly certain to the species Acrodus minimus, they are quite similar in size and measurement. However, there are a series of more round ones, more frequent than the elongated ones. |
| |
|
|
|
A marine mackerel shark, type member of the family Agaleidae inside Euselachii. It was assigned to the genus Otodus. Very unlikely to be the genus Otodus, since its lineage appears on the Late Cretaceous. Some small teeth not exceeding 7 mm in height, garish smooth, that resembles strongly those found in the Cretaceous Otodus-like teeth.Was later confirmed to be teeth from Agaleus. |
||


Actinopteri
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Teeth |
A marine Osteichthyes, the youngest representative of the family Saurichthyidae inside Chondrostei. Was identified as Saurichthys. Of this characteristic Genus were found 3 teeth, resembling superficially Saurichthys acuminata, and a third that looks more like belonging to Saurichthys longidens. |
Saurorhynchus a coeval genus | |
References
- Michelsen, O., Nielsen, L. H., Johannessen, P. N., Andsbjerg, J., & Surlyk, F. (2003). Jurassic lithostratigraphy and stratigraphic development onshore and offshore Denmark. Geological Survey of Denmark and Greenland (GEUS) Bulletin, 1, 145-216.
- Gry, H., Jørgart, T., & Poulsen, V. (1969). Geologi pa Bornholm. Mineralogisk Mus.
- Gravesen, P. (1982). lithostratigraphy and sedimentary evolution of the Triassic, Jurassic and Lower Cretaceous of Bornholm, Denmark.
- Gry, H. E. L. G. E. (1969). Megaspores from the Jurassic of the island of Bornholm, Denmark. Meddelelser fra Dansk Geologisk Forening, 19, 69-89.
- Bergelin, I., Obst, K., Söderlund, U., Larsson, K., & Johansson, L. (2011). Mesozoic rift magmatism in the North Sea region: 40 Ar/39 Ar geochronology of Scanian basalts and geochemical constraints. International Journal of Earth Sciences, 100(4), 787-804.
- Nielsen, L.H. 1987: Progress report 1.1.1988. Biostratigraphy and organic geochemistry of the Mesozoic on Bornholm. DGU 215 confidential report 31, 35 pp. Copenhagen: Geological Survey of Denmark.
- Koppelhus, E.B. 1988: Catalogue of spores and pollen from the Lower–Middle Jurassic Bagå Formation on Bornholm, Denmark. DGU confidential report 21, 42 pp. Copenhagen: Geological 214 Survey of Denmark
- Koppelhus, E.B. & Batten, D.J. 1992: Megaspore assemblages from the Jurassic and lowermost Cretaceous of Bornholm, Denmark. Danmarks Geologiske Undersøgelse Serie A 32, 81 pp
- Zimmermann, J., Franz, M., Schaller, A., & Wolfgramm, M. (2017). The Toarcian-Bajocian deltaic system in the North German Basin: Subsurface mapping of ancient deltas-morphology, evolution and controls. Sedimentology, 65(3), 897–930. doi:10.1111/sed.12410
- Barth, G., Pieńkowski, G., Zimmermann, J., Franz, M., & Kuhlmann, G. (2018). Palaeogeographical evolution of the Lower Jurassic: high-resolution biostratigraphy and sequence stratigraphy in the Central European Basin. Geological Society, London, Special Publications, 469(1), 341–369.
- Bromley, R. G., & Uchman, A. (2003). Trace fossils from the Lower and Middle Jurassic marginal marine deposits of the Sorthat Formation, Bornholm, Denmark. Bulletin of the Geological Society of Denmark, 52, 185-208.
- Hoelstad,T. (1985) Palynology of the Uppermost Lower to Middle Jurassic strata on Bornholm, Denmark. Geological Society of Denmark, Bulletin (Meddelelser Fra Dansk Geologisk Forening) Vol. 34 P. 111- 132
- PETERSEN, H.I., NIELSEN, L.H., KOPPELHUS, E.B., and SØRENSEN, H.S., 2003, Early and Middle Jurassic mires of Bornholm and the Fennoscandian Border Zone: A comparison of depositional environments and vegetation: Geological Survey of Denmark and Greenland Bulletin, v. 1, p. 631–656
- Zimmermann, J., Franz, M., Heunisch, C., Luppold, F. W., Mönnig, E., & Wolfgramm, M. (2015). Sequence stratigraphic framework of the Lower and Middle Jurassic in the North German Basin: epicontinental sequences controlled by Boreal cycles. Palaeogeography, Palaeoclimatology, Palaeoecology, 440, 395-416.
- O. Graversen: Upper Triassic–Lower Cretaceous seismic sequence stratigraphy and basin tectonics at Bornholm, Denmark, Tornquist Zone, NW Europe Mar. Pet. Geol., 21 (2004), pp. 579-612
- Koppelhus, E. B., & Nielsen, L. H. (1994). Palynostratigraphy and palaeoenvironments of the lower to middle jurassic bagå formation of bornholm, denmark. Palynology, 18(1), 139–194. doi:10.1080/01916122.1994.9989443
- Nielsen, L.H. (1988): Lithostratigraphy and depositional environment of the Lower and Middle Jurassic sequence drilled in shallow wells. Geological Survey of Denmark, Report no. 24, 48 pgs
- Petersen, H. I. (1993). Petrographic facies analysis of Lower and Middle Jurassic coal seams on the island of Bornholm, Denmark. International Journal of Coal Geology, 22(3-4), 189-216.
- Petersen, H. I., Nielsen, L. H., Koppelhus, E. B., & Sørensen, H. S. (2003). Early and Middle Jurassic mires of Bornholm and the Fennoscandian Border Zone: a comparison of depositional environments and vegetation. Geological Survey of Denmark and Greenland (GEUS) Bulletin, 1, 631-656.
 This article incorporates text available under the CC BY 4.0 license.
This article incorporates text available under the CC BY 4.0 license. - Crawford, A. J. (2015). Understanding fire histories: the importance of charcoal morphology.
- Sachs, S., Hornung, J. J., Lierl, H.-J., & Kear, B. P. (2016). Plesiosaurian fossils from Baltic glacial erratics: evidence of Early Jurassic marine amniotes from the southwestern margin of Fennoscandia. Geological Society, London, Special Publications, 434(1), 149–163. doi:10.1144/sp434.14
- Petersen,H.I. et al. (1995) Controls on Peat accumulation and depositional environments of a Coal-bearing Coastal Plain succession of a Pull-Apart Basin; a petrographic, geochemical and sedimentological study, Lower Jurassic, Denmark International Journal of Coal Geology Vol. 27 # 2 P. 99- 129
- Seidenkrantz,M.S. et al. (1993) Biostratigraphy and palaeoenvironmental analysis of a Lower to Middle Jurassic succession on Anholt, Denmark. Journal of Micropalaeontology Vol. 12 # 2 P. 201- 218
- Wade-Murphy, J., Kuerschner, W. M., & Hesselbo, S. P. (2006). Early Toarcian vegetation history from the Korsodde Section of Bornholm (Denmark) and its possible impact on terrestrial carbon isotope records. In 7th European Palaeobotany Palynology Conference, Prague (p. 153).
- Hesselbo, S. P., Gröcke, D. R., Jenkyns, H. C., Bjerrum, C. J., Farrimond, P., Bell, H. S. M., & Green, O. R. (2000). Massive dissociation of gas hydrate during a Jurassic oceanic anoxic event. Nature, 406(6794), 392-395.
- Michelsen,O. (1979) Report on the Jurassic of the Hobro No.1 and Voldum No.1 Borings, Denmark. Danmarks Geologiske Undersиgelse P. 141- 149
- Koppelhus,E.B. et al. (1996) Application of a palynomorph zonation to a series of short borehole sections, Lower to Middle Jurassic, Oresund, Denmark. (Principles and Applications. Applications. J.Jansonius and D.C.McGregor, editors.) American Association of Stratigraphic Palynologists:Principles and Applications. Applications (J.Jansonius and D.C.McGregor,Editors) Vol. 2 P. 779- 793
- Tekleva, M. V., Krassilov, V. A., Kvacek, J., & Van Konijnenburg-van Cittert, J. H. A. (2006). Pollen genus Eucommiidites: ultrastructure and affinities. ACTA PALAEOBOTANICA-KRAKOW-, 46(2), 137.
- Crane, P. R. (1996). The fossil history of the Gnetales. International Journal of Plant Sciences, 157(S6), S50-S57.
- Rydin, C., Pedersen, K. R., Crane, P. R., & Friis, E. M. (2006). Former diversity of Ephedra (Gnetales): evidence from early Cretaceous seeds from Portugal and North America. Annals of Botany, 98(1), 123-140.
- Wade-Murphy, J., Kuerschner, W. M., (2006). A new technique to infer the botanical affinity of palynomorphs, and its application on spheripollenites psilatus from the toarcian of bornholm, denmark. In 7 th European Palaeobotany Palynology Conference, Prague (p. 153).
- Wade-Murphy, Kuerschner, W. M., & Hesselbo, S. P. (2006): Abrupt and gradual vegetation changes associated with Toarcian global change inferred from high resolution palynological study of the Korsodde section on Bornholm (dK). In Geophysical Research Abstracts (Vol. 8, p. 00357).
- Bartholin, C.T. (1892): Nogle i den bornholmske Juraformation forekommende Planteforsteninger. Botanisk Tidsskrift, 18: 12-28.
- Bartholin, C.T. (1894): Nogle i den bornholmske Juraformation forekommende Planteforsteninger. Botanisk Tidsskrift, 19:87-115.
- Hjort, A. (1899): Om Vellengsbyleret og dets Flora. Danmarks Geologiske Undersøgelse, II rk. 10: 61-86.
- Möller, H., 1903, Bidrag till Bornholms fossila flora (Rhät och Lias)-Gymnospermer: Kungliga Svenska Vetenskapsakademiens Handlingar, v. 36(6), p. 3-48.
- Möller, H. (1902). Bidrag till Bornholms fossila flora: Pteridofyter (Vol. 38, No. 5). E. Malmströms boktryckeri.
- Mehlqvist, K., Vajda, V., & Larsson, L. M. (2009). A Jurassic (Pliensbachian) flora from Bornholm, Denmark–a study of a historic plant-fossil collection at Lund University, Sweden. GFF, 131(1-2), 137-146.
- McElwain, J. C., Wade-Murphy, J., & Hesselbo, S. P. (2005). Changes in carbon dioxide during an oceanic anoxic event linked to intrusion into Gondwana coals. Nature, 435(7041), 479-482.
- Seward, A. C. (1903). On the occurrence of Dictyozamites in England, with remarks on European and eastern mesozoic floras. Quarterly Journal of the Geological Society, 59(1-4), 217-233.
- Florin, R. (1933). Über Nilssoniopteris glandulosa n. sp., eine Bennettitacee aus der Juraformation Bornholms. Arkiv för Botanik, 25(20), 19.
- Florin, R. (1958): On Jurassic taxads and conifers from north-western Europe and eastern Greenland. Acta Horti Bergiani, 17: 402 pgs.
- Yao, X., Zhou, Z., & Zhang, B. (1998). Reconstruction of the Jurassic conifer Sewardiodendron laxum (Taxodiaceae). American journal of botany, 85(9), 1289-1300.
- Keighley, D. G., & Pickerill, R. K. (1995). Commentary: the ichnotaxa Palaeophycus and Planolites: historical perspectives and recommendations.
- Bandel, K. (1973). A new name for the ichnogenus Cylindrichnus Bandel, 1967. Journal of Paleontology, 1002-1002.
- Knaust, D. (2018). The ichnogenus Teichichnus Seilacher, 1955. Earth-Science Reviews, 177, 386-403.
- Osgood, R. G. (1975). The history of invertebrate ichnology. In The study of trace fossils (pp. 3–12). Springer, Berlin, Heidelberg.
- Hertweck, G., Wehrmann, A., & Liebezeit, G. (2007). Bioturbation structures of polychaetes in modern shallow marine environments and their analogues to Chondrites group traces. Palaeogeography, Palaeoclimatology, Palaeoecology, 245(3-4), 382–389.
- Chakraborty, A., & Bhattacharya, H. N. (2013). Spreiten burrows: a model based study on Diplocraterion parallelum. Geospectrum, 296-299.
- Ekdale, A. A., & Lewis, D. W. (1991). Trace fossils and paleoenvironmental control of ichnofacies in a late Quaternary gravel and loess fan delta complex, New Zealand. Palaeogeography, Palaeoclimatology, Palaeoecology, 81(3-4), 253–279. doi:10.1016/0031-0182(91)90150-p
- Malling, C., & Grönwall, K. A. (1912). En fauna i Bornholms Lias. Geologiska Föreningen i Stockholm Förhandlingar, 34(3), 366-367.
- Åhlander, F. E. (1919). Förteckning öfver svensk geologisk, paleontologisk, petrografisk och mineralogisk litteratur för åren 1907–1917. Geologiska Föreningen i Stockholm Förhandlingar, 41(7), 539-606.
- W. Ohmert, V. Allia C. Arias et al. 1996. Die Grenzziehung Unter-/Mitteljura (Toarcium/Aalenium) bei Wittnau und Fuentelsaz [Determinig the boundary between Toarcium/Aalenium near Wittnau and Fuentelsaz]. Informationen Geologisches Landesamt Baden-Württemberg 8:1–52
- F. Papier. 2001. Die Gundershoffener Klamm. Fossilien 6/01:368–374
- Beurlen, K. (1932). Brachyurenreste aus dem Lias von Bornholm mit Beiträgen zur Phylogenie und Systematik der Brachyuren Decapoden. Paläontologische Zeitschrift, 14(1-2), 52–66. doi:10.1007/bf03041617
- Schweigert G. (2012): Verschollene Lias-Krabben von Bornholm – ein Paläokrimi (noch) ohne Happyend. Fossilien 2012: 301-304.
- Schweitzer, C. E., & Feldmann, R. M. (2007). A new Classification for some Jurassic Brachyura (Crustacea: Decapoda: Brachyura: Homolodromioidea): families GoniodromitidaeBeurlen, 1932 and Tanidromitidae new family. Senckenbergiana Lethaea, 87(2), 119–155. doi:10.1007/bf03043911
- Duffin, C.J. & Ward, D.J. 1983. Teeth of a new Neoselachian shark from the British Lower Jurassic. -Palaeontology 26, 839-844.