Embolomeri
Embolomeri is an order of tetrapods or stem-tetrapods, possibly members of Reptiliomorpha. Embolomeres first evolved from reptile-like amphibians in the Early Carboniferous (Mississippian). They were strongly specialized semiaquatic predators with long bodies for eel-like undulating swimming. Embolomeres are characterized by their vertebral centra, which are formed by two cylindrical segments, the pleurocentrum at the rear and intercentrum at the front. These segments are equal in size. Most other tetrapods have pleurocentra and intercentra which are drastically different in size and shape.[1]
| Embolomeres | |
|---|---|
 | |
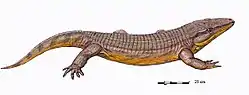
| Restoration of Archeria from the Lower Permian of Texas. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Superclass: | Tetrapoda |
| Clade: | Reptiliomorpha (?) |
| Order: | †Embolomeri Cope, 1885 |
| Subgroups | |
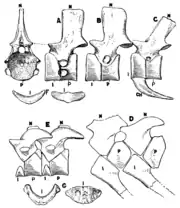
Embolomeres were among the earliest large carnivorous tetrapods, with members such as the crocodilian-like Proterogyrinus appearing in the mid-Carboniferous period. However, they declined in diversity during the Permian period, although the most common embolomere known, Archeria, did live during this time. Embolomeres went extinct shortly before the end of the Permian.
The order Embolomeri was first named by Edward Drinker Cope in 1884 during his revision of "batrachian" (amphibian) evolution. Embolomeri was differentiated from several other newly named amphibian orders, such as "Rachitomi", by the presence of intercentra and pleurocentra of the same size and shape, that being large cylinders. At the time, embolomere fossils were uncommon, so Cope could only identify "cricotids" such as Cricotus as possessing embolomerous vertebrae.[1] The genus name "Cricotus" is dubious, as it has been used by Cope to refer to embolomere fossils spanning anywhere between mid-Pennsylvanian deposits of Illinois and the Permian red beds of Texas. Most paleontologists now refer the red bed "Cricotus" specimens to the genus Archeria.[2]
Michel Laurin (1998) formally defined Embolomeri as "the last common ancestor of Proterogyrinus and Archeria and all of its descendants."[3] This definition excludes Eoherpeton, which is almost always considered a close ally of the group. Some authors describe an "embolomere clade" containing not only the taxa included by Laurin, but also Eoherpeton and occasionally even chroniosuchians and Silvanerpeton.[4][5][6]
Classification
Like most early tetrapod or stem-tetrapod groups, the phylogenetic position of embolomeres is controversial. For much of the later 20th century, they were placed in the group Anthracosauria, a collection of tetrapods which resembled the amphibians biologically but were probably the ancestors of amniotes (reptiles and synapsids) rather than modern lissamphibians. Lissamphibians are often considered to be descendants of the temnospondyls, an entirely different group of crocodile-like amphibious tetrapods.[7]
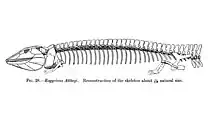
Anthracosauria is sometimes considered synonymous with Embolomeri, and the group's namesake, Anthracosaurus, is an embolomere. However, other authors use the term "Anthracosauria" in reference to a broader group including embolomeres in combination with various other reptile-like amphibians (reptiliomorphs). Many studies conducted since the 1990s have also placed the group Lepospondyli as closer to amniotes than embolomeres were. Lepospondyls are a particularly unusual group of tetrapods, with some members (i.e. brachystelechids) very similar to lissamphibians and others (i.e. tuditanids) very similar to amniotes. If lepospondyls are both close relatives of amniotes and the ancestors of modern amphibians, then that means that crown-Tetrapoda (descendants of the common ancestor to all living tetrapods) is a much more restricted group than previously assumed. In this situation, various traditional orders of Tetrapoda such as Embolomeri and Temnospondyli actually would qualify as stem-tetrapods due to having evolved prior to the split between modern amphibians and amniotes.[8]
On the other hand, if temnospondyls are the ancestors of modern amphibians, then embolomeres would likely be reptiliomorphs, part of Tetrapoda.[7] However, even this classification is not stable, as some analyses have found embolomeres to be more basal than temnospondyls.[9] Chroniosuchians are thought to be close relatives, but are generally agreed to lie outside Embolomeri proper.[10]
Below is a cladogram from Ruta et al. (2003):[7]
| Tetrapoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
List of genera
| Name | Year | Age | Location | Notes | Images |
|---|---|---|---|---|---|
| Anthracosaurus | 1863 | Pennsylvanian (Westphalian A-B) | The namesake of Anthracosauria, possessed a small number of large fangs compared to other embolomeres |  | |
| Archeria | 1918 | Early Permian | A common, late-surviving member of the group, sometimes considered a species of Cricotus |  | |
| Aversor | 1985 | Early Permian (Ufimian) | Supposedly the latest surviving eogyrinid, but very poorly known | ||
| Calligenethlon | 1934 | Pennsylvanian (Bashkirian) | The largest named animal known from the Joggins Fossil Cliffs | ||
| Cricotus | 1875 | Pennsylvanian | Responsible for the identification of Embolomeri as a unique order of tetrapods, although its history is convoluted and its taxonomic validity is questionable | ||
| Carbonoherpeton | 1985 | Pennsylvanian (Westphalian D) | A lightly built member of the group with characteristics of both archeriids and eogyrinids | ||
| Diplovertebron | 1879 | Pennsylvanian (Moscovian) | A small member of the group sometimes confused with Gephyrostegus |  | |
| Eobaphetes | 1916 | Pennsylvanian | A possible relative of Anthracosaurus, originally named "Erpetosuchus" until it was determined that the name was preoccupied by a Triassic reptile. | ||
| Eogyrinus | 1926 | Pennsylvanian (Westphalian B) | One of the largest and most well-known of the eogyrinids, although sometimes synonymized with Pholiderpeton |  | |
| Eoherpeton | 1975 | Pennsylvanian (Visean-Bashkirian) | One of the oldest and most basal members of the group (if it even counts as part of it), without vertebrae that were not fully embolomerous |  | |
| Leptophractus | 1873 | Pennsylvanian (Westphalian D) | Known from a skull found at the Linton Diamond Mine | ||
| Neopteroplax | 1963 | Pennsylvanian | One of the largest Carboniferous limbed vertebrates known from North America | _(Birmingham_Shale%252C_Upper_Pennsylvanian%253B_Bloomingdale%252C_Ohio%252C_USA)_1_(49761151626).jpg.webp) | |
| Palaeoherpeton | 1970 | Pennsylvanian (Westphalian A-B) | An eogyrinid known as Palaeogyrinus from 1926 to 1970, until it was determined that this name was occupied by a genus of beetles | ||
|
Papposaurus |
1914 | Mississippian-Pennsylvanian (Namurian) | Known from a femur, may have been a relative of Proterogyrinus | ||
| Pholiderpeton | 1869 | Pennsylvanian (Westphalian A-B) | An eogyrinid similar to Eogyrinus which would take priority if the two were synonymized |  | |
| Proterogyrinus | 1970 | Mississippian (Serpukhovian) | An early member of the group possessing robust limbs but lacking certain adaptations of later members of the group |  | |
| Pteroplax | 1868 | Pennsylvanian (Westphalian B) | Poorly known despite being among the first embolomeres to be described |  | |
| Seroherpeton | 2020 | Late Permian (Wuchiapingian) | The youngest known embolomere | ||
| Spondylerpeton | 1912 | Pennsylvanian | A close relative of Cricotus known from vertebrae found at the Mazon Creek fossil beds | ||
References
- Cope, E.D. (January 1884). "The Batrachia of the Permian Period of North America". The American Naturalist. 18: 26–39. doi:10.1086/273563.
- Romer, Alfred Sherwood (11 January 1957). "The Appendicular Skeleton of the Permian Embolomerous Amphibian Archeria" (PDF). Contributions from the Museum of Paleontology. 13 (5): 105–159.
- Laurin, Michel (1998). "The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I. Systematics, middle ear evolution and jaw suspension". Annales des Sciences Naturelles - Zoologie et Biologie Animale. 19 (1): 1–42. doi:10.1016/S0003-4339(98)80132-9.
- Klembara, Jozef; Clack, Jennifer A.; Čerňanský, Andrej (2010-09-16). "The anatomy of palate of Chroniosaurus dongusensis (Chroniosuchia, Chroniosuchidae) from the Upper Permian of Russia". Palaeontology. 53 (5): 1147–1153. doi:10.1111/j.1475-4983.2010.00999.x. ISSN 0031-0239.
- Carroll, Robert L. (2012-10-18). "The Importance of Recognizing Our Limited Knowledge of the Fossil Record in the Analysis of Phylogenetic Relationships among Early Tetrapods". Fieldiana Life and Earth Sciences. 5: 5–16. doi:10.3158/2158-5520-5.1.5. ISSN 2158-5520.
- Sookias, Roland B.; Böhmer, Christine; Clack, Jennifer A. (2014-10-07). "Redescription and Phylogenetic Analysis of the Mandible of an Enigmatic Pennsylvanian (Late Carboniferous) Tetrapod from Nova Scotia, and the Lability of Meckelian Jaw Ossification". PLoS ONE. 9 (10): e109717. doi:10.1371/journal.pone.0109717. ISSN 1932-6203. PMC 4188710. PMID 25290449.
- Marcello Ruta, Michael I. Coates and Donald L. J. Quicke (2003). "Early tetrapod relationships revisited" (PDF). Biological Reviews. 78 (2): 251–345. doi:10.1017/S1464793102006103. PMID 12803423.
- Laurin, M.; Reisz, R.R. (1999). "A new study of Solenodonsaurus janenschi, and a reconsideration of amniote origins and stegocephalian evolution". Canadian Journal of Earth Sciences. 36 (8): 1239–1255. doi:10.1139/e99-036.
- Clack, J. A. (July 2002). "An early tetrapod from 'Romer's Gap'". Nature. 418 (6893): 72–76. doi:10.1038/nature00824. ISSN 0028-0836. PMID 12097908.
- Chen, Jianye; Liu, Jun (2020-12-01). "The youngest occurrence of embolomeres (Tetrapoda: Anthracosauria) from the Sunjiagou Formation (Lopingian, Permian) of North China". Fossil Record. 23 (2): 205–213. doi:10.5194/fr-23-205-2020. ISSN 2193-0066.