Gnetophyta
Gnetophyta (/nɛˈtɒfɪtə, ˈnɛtoʊfaɪtə/) is a division of plants, grouped within the gymnosperms (which also includes conifers, cycads, and ginkgos), that consists of some 70 species across the three relict genera: Gnetum (family Gnetaceae), Welwitschia (family Welwitschiaceae), and Ephedra (family Ephedraceae). Fossilized pollen attributed to a close relative of Ephedra has been dated as far back as the Early Cretaceous.[1] Though diverse in the Early Cretaceous, only three families, each containing a single genus, are still alive today. The primary difference between gnetophytes and other gymnosperms is the presence of vessel elements, a system of conduits that transport water within the plant, similar to those found in flowering plants. Because of this, gnetophytes were once thought to be the closest gymnosperm relatives to flowering plants, but more recent molecular studies have brought this hypothesis into question.
| Gnetophyta | |
|---|---|
 | |
| Welwitschia mirabilis female plant with cones | |
| Scientific classification | |
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Division: | Gnetophyta Bessey 1907 |
| Class: | Gnetopsida Thom 1886 |
| Families & Genera | |
|
Gnetaceae | |
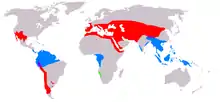 | |
| Distribution, separated by genus: Green – Welwitschia Blue – Gnetum Red – Ephedra Purple – Gnetum and Ephedra | |
Though it is clear they are all closely related, the exact evolutionary inter-relationships between gnetophytes are unclear. Some classifications hold that all three genera should be placed in a single order (Gnetales), while other classifications say they should be distributed among three separate orders, each containing a single family and genus. Most morphological and molecular studies confirm that the genera Gnetum and Welwitschia diverged from each other more recently than they did from Ephedra.[2][3][4][5][6]

_1.jpg.webp)
.jpg.webp)



Ecology and morphology
Unlike most biological groupings, it is difficult to find many common characteristics between all of the members of the gnetophytes.[7] The two common characteristics most commonly used are the presence of enveloping bracts around both the ovules and microsporangia as well as a micropylar projection of the outer membrane of the ovule that produces a pollination droplet,[8] though these are highly specific compared to the similarities between most other plant divisions. L. M. Bowe refers to the gnetophyte genera as a "bizarre and enigmatic" trio[3] because, the gnetophytes' specialization to their respective environments is so complete that they hardly resemble each other at all. Gnetum species are mostly woody vines in tropical forests, though the best-known member of this group, Gnetum gnemon,[9] is a tree native to western Malesia. The one remaining species of Welwitschia, Welwitschia mirabilis, native only to the dry deserts of Namibia and Angola, is a ground-hugging species with only two large strap-like leaves that grow continuously from the base throughout the plant's life. Ephedra species, known as "jointfirs" in the United States, have long slender branches which bear tiny scale-like leaves at their nodes. Infusions from these plants have been traditionally used as a stimulant, but ephedrine is a controlled substance today in many places because of the risk of harmful or even fatal overdosing.
Fossil Gnetophyta
Knowledge of gnetophyte history through fossil discovery has increased greatly since the 1980s.[2] Gnetophyte fossils have been found that date from the Permian[10] and the Triassic. Fossils dating back to the Jurassic have been found, though whether or not they belong to the gnetophytes is uncertain.[11] Overall, the fossil record is richest in the early Cretaceous, with fossils of plants, seeds, and pollen have been found that can clearly be assigned to the gnetophytes.[11]
Classification
With just three well-defined genera within an entire division, there still is understandable difficulty in establishing an unambiguous interrelationship among them; in earlier times matters were even more difficult and we find for example Pearson in the early 20th century speaking of the class Gnetales, rather than the order.[12] G. H. M. Lawrence referred to them as an order, but remarked that the three families were distinct enough to deserve recognition as separate orders.[13] Foster & Gifford accepted this principle, and placed the three orders together in a common class for convenience, which they called Gnetopsida.[14] In general the evolutionary relationships among the seed plants still are unresolved, and the Gnetophyta have played an important role in the formation of phylogenetic hypotheses. Molecular phylogenies of extant gymnosperms have conflicted with morphological characters with regard to whether the gymnosperms as a whole (including gnetophytes) comprise a monophyletic group or a paraphyletic one that gave rise to angiosperms. At issue is whether the Gnetophyta are the sister group of angiosperms, or whether they are sister to, or nested within, other extant gymnosperms. Numerous fossil gymnosperm clades once existed that are morphologically at least as distinctive as the four living gymnosperm groups, such as Bennettitales, Caytonia and the glossopterids. When these gymnosperm fossils are considered, the question of gnetophyte relationships to other seed plants becomes even more complicated. Several hypotheses, illustrated below, have been presented to explain seed plant evolution. Morphological studies have supported a close relationship between Gnetophyta, Bennettitales and the Erdtmanithecales.[15]
Recent research by Lee EK, Cibrian-Jaramillo A, et al. (2011) suggests that the Gnetophyta are a sister group to the rest of the gymnosperms,[16] contradicting the anthophyte hypothesis, which held that gnetophytes were sister to the flowering plants.
Anthophyte hypothesis
From the early twentieth century, the anthophyte hypothesis was the prevailing explanation for seed plant evolution, based on shared morphological characters between the gnetophytes and angiosperms. In this hypothesis, the gnetophytes, along with the extinct order Bennettitales, are sister to the angiosperms, forming the "anthophytes".[8] Some morphological characters that were suggested to unite the anthophytes include vessels in wood, net-veined leaves (in Gnetum only), lignin chemistry, the layering of cells in the apical meristem, pollen and megaspore features (including thin megaspore wall), short cambial initials, and lignin syringal groups.[8][17][18][19] However, most genetic studies, as well as more recent morphological analyses,[20] have rejected the anthophyte hypothesis.[3][21][22][23][24][25][26][27][28][29] Several of these studies have suggested that the gnetophytes and angiosperms have independently derived characters, including flower-like reproductive structures and tracheid vessel elements, that appear shared but are actually the result of parallel evolution.[3][8][22]
|
Ginkgo | |||||||
|
cycads | |||||||
|
conifers | |||||||
| anthophytes |
| ||||||
Gnetifer hypothesis
In the gnetifer hypothesis, the gnetophytes are sister to the conifers, and the gymnosperms are a monophyletic group, sister to the angiosperms. The gnetifer hypothesis first emerged formally in the mid-twentieth century, when vessel elements in the gnetophytes were interpreted as being derived from tracheids with circular bordered pits, as in conifers.[8] It did not gain strong support, however, until the emergence of molecular data in the late 1990s.[21][27][30][31] Although the most salient morphological evidence still largely supports the anthophyte hypothesis, there are some more obscure morphological commonalities between the gnetophytes and conifers that lend support to the gnetifer hypothesis. These shared traits include: tracheids with scalariform pits with tori interspersed with annular thickenings, absence of scalariform pitting in primary xylem, scale-like and strap-shaped leaves of Ephedra and Welwitschia; and reduced sporophylls.[26][29][32]
|
angiosperms (flowering plants) | ||||||||||||||||
| gymnosperms |
| |||||||||||||||
Gnepine hypothesis
The gnepine hypothesis is a modification of the gnetifer hypothesis, and suggests that the gnetophytes belong within the conifers as a sister group to the Pinaceae.[8] According to this hypothesis, the conifers as currently defined are not a monophyletic group, in contrast with molecular findings that support its monophyly.[30] All existing evidence for this hypothesis comes from molecular studies since 1999.[3][4][22][24][26][27][29][32] However, the morphological evidence remains difficult to reconcile with the gnepine hypothesis. If the gnetophytes are nested within conifers, they must have lost several shared derived characters of the conifers (or these characters must have evolved in parallel in the other two conifer lineages): narrowly triangular leaves (gnetophytes have diverse leaf shapes), resin canals, a tiered proembryo, and flat woody ovuliferous cone scales.[26] These kinds of major morphological changes are not without precedent in the Pinales, however: the Taxaceae, for example, have lost the classical cone of the conifers in favor of a single-terminal ovule surrounded by a fleshy aril.[22]
|
angiosperms (flowering plants) | ||||||||||||||||||||||
| gymnosperms |
| |||||||||||||||||||||
Gnetophyte-sister hypothesis
Some partitions of the genetic data suggest that the gnetophytes are sister to all of the other extant seed plant groups.[5][8][26][29][30][33][34] However, there is no morphological evidence nor examples from the fossil record to support the gnetophyte-sister hypotheses.[32]
| ||||||||||||||||
Fossil lineages
- Ephedraceae
- Leongathia V.A. Krassilov ,D.L. Dilcher & J.G. Douglas 1998[35] Koonwarra fossil bed, Australia, Aptian
- Jianchangia Yang, Wang and Ferguson, 2020[36] Jiufotang Formation, China, Aptian
- Eamesia Yang, Lin and Ferguson, 2018[37] Yixian Formation, China, Aptian
- Prognetella Krassilov et Bugdaeva, 1999 Yixian Formation, China, Aptian (initially interpreted as an angiosperm)[38]
- Chengia Yang, Lin & Wang, 2013,[39] Yixian Formation, China, Aptian
- Gnetaceae
- Khitania Guo et al. 2009[40] Yixian Formation, China, Aptian
- Welwitschiaceae
- Priscowelwitschia
- Incertae sedis:
- Drewria Crane & Upchurch, 1987 Potomac Group, USA, Albian (possible affinities to Welwitschiaceae)[41]
- Bicatia Friis, Pedersen and Crane, 2014[41] Figueira da Foz Formation, Portugal, late Aptian early Albian, Potomac Group, USA, Albian (possible affinities to Welwitschiaceae)
- Liaoningia Yang et al, 2017[42] Yixian Formation, China, Aptian
- Protognetum Y. Yang, L. Xie et D.K. Ferguson, 2017[43] Daohugou, China, Callovian
- Itajuba Ricardi-Branco et al, 2013,[44] Crato Formation, Brazil, Aptian
- Protoephedrites Rothwell et Stockey, 2013[45] Canada, Valanginian (possible ephedroid affinities)
- Siphonospermum Rydin et Friis, 2010[46] Yixian Formation, China, Aptian
References
| Wikimedia Commons has media related to Gnetophyta. |
- "Morphology and affinities of an Early Cretaceous Ephedra".
- Peter R. Crane; Patrick Herendeen; Else Marie Friis (2004). "Fossils and plant phylogeny". American Journal of Botany. 91 (10): 1683–1699. doi:10.3732/ajb.91.10.1683. PMID 21652317.
- Bowe, L.M.; Coat, G.; dePamphilis, C.W. (2000). "Phylogeny of seed plants based on all three genomic compartments: Extant gymnosperms are monophyletic and Gnetales' closest relatives are conifers". Proceedings of the National Academy of Sciences. 97 (8): 4092–4097. Bibcode:2000PNAS...97.4092B. doi:10.1073/pnas.97.8.4092. PMC 18159. PMID 10760278.
- Gugerli, F.; Sperisen, C.; Buchler, U.; Brunner, L.; Brodbeck, S.; Palmer, J.D.; Qiu, Y.L. (2001). "The evolutionary split of Pinaceae from other conifers: evidence from an intron loss and a multigene phylogeny". Molecular Phylogenetics and Evolution. 21 (2): 167–175. doi:10.1006/mpev.2001.1004. PMID 11697913.
- Rai, H.S.; Reeves, P.A.; Peakall, R.; Olmstead, R.G.; Graham, S.W. (2008). "Inference of higher-order conifer relationships from a multi-locus plastid data set". Botany. 86 (7): 658–669. doi:10.1139/B08-062.
- Ickert-Bond, S. M.; C. Rydin & S. S. Renner (2009). "A fossil-calibrated relaxed clock for Ephedra indicates an Oligocene age for the divergence of Asian and New World clades, and Miocene dispersal into South America" (PDF). Journal of Systematics and Evolution. 47 (5): 444–456. doi:10.1111/j.1759-6831.2009.00053.x. S2CID 55148071.
- Arber, E.A.N.; Parkin, J. (1908). "Studies on the evolution of the angiosperms: the relationship of the angiosperms to the Gnetales". Annals of Botany. 22 (3): 489–515. doi:10.1093/oxfordjournals.aob.a089185.
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F.; and Donoghue, M.J. (2008) Plant Systematics: A Phylogenetics Approach. 3rd ed. Sunderland, Massachusetts, USA: Sinauer Associates, Inc.
- Wan T, Liu Z M, Li L F, et al. A genome for gnetophytes and early evolution of seed plants[J]. Nature plants, 2018, 4(2): 82.
- Zi-Qiang Wang (2004). "A New Permian Gnetalean Cone as Fossil Evidence for Supporting Current Molecular Phylogeny". Annals of Botany. 94 (2): 281–288. doi:10.1093/aob/mch138. PMC 4242163. PMID 15229124.
- Catarina Rydin; Kaj Raunsgaard Pedersen; Peter R. Crane; Else Marie Friis (2006). "Former Diversity of Ephedra (Gnetales): Evidence from Early Cretaceous Seeds from Portugal and North America". Annals of Botany. 98 (1): 123–140. doi:10.1093/aob/mcl078. PMC 2803531. PMID 16675607.
- Pearson, H. H. W. Gnetales. Cambridge University Press 1929. Reissued 2010. ISBN 978-1108013987
- Lawrence, George Hill Mathewson. Taxonomy of vascular plants. Macmillan, 1951
- Foster, Adriance S., Gifford, Ernest M. Jr. Comparative Morphology of Vascular Plants Freeman 1974. ISBN 0-7167-0712-8
- Friis, Else Marie; Crane, Peter R.; Pedersen, Kaj Raunsgaard; Bengtson, Stefan; Donoghue, Philip C. J.; Grimm, Guido W.; Stampanoni, Marco (November 2007). "Phase-contrast X-ray microtomography links Cretaceous seeds with Gnetales and Bennettitales". Nature. 450 (7169): 549–552. Bibcode:2007Natur.450..549F. doi:10.1038/nature06278. ISSN 0028-0836. PMID 18033296. S2CID 1198220.
- Lee EK, Cibrian-Jaramillo A, Kolokotronis SO, Katari MS, Stamatakis A, et al. (2011). "A Functional Phylogenomic View of the Seed Plants". PLOS Genet. 7 (12): e1002411. doi:10.1371/journal.pgen.1002411. PMC 3240601. PMID 22194700.
- Donoghue, M.J.; Doyle, J.A. (2000). "Seed plant phylogeny: demise of the anthophyte hypothesis?". Current Biology. 10 (3): R106–R109. doi:10.1016/S0960-9822(00)00304-3. PMID 10679315. S2CID 16558206.
- Loconte, H.; Stevenson, D.W. (1990). "Cladistics of the Spermatophyta". Brittonia. 42 (3): 197–211. doi:10.2307/2807216. JSTOR 2807216. S2CID 36908568.
- Nixon, K.C.; Crepet, W.L.; Stevenson, D.; Friis, E.M. (1994). "A reevaluation of seed plant phylogeny". Annals of the Missouri Botanical Garden. 81 (3): 494–533. doi:10.2307/2399901. JSTOR 2399901.
- Coiro, M.; Chomicki, G.; Doyle, J.A. (2018). "Experimental signal dissection and method sensitivity analyses reaffirm the potential of fossils and morphology in the resolution of the relationship of angiosperms and Gnetales". Paleobiology. 44 (3): 490–510. doi:10.1017/pab.2018.23. S2CID 91488394.
- Chaw, S.M.; Aharkikh, A.; Sung, H.M.; Lau, T.C.; Li, W.H. (1997). "Molecular phylogeny of extant gymnosperms and seed plant evolution: analysis of nuclear 18S rRNA sequences". Molecular Biology and Evolution. 14 (1): 56–68. doi:10.1093/oxfordjournals.molbev.a025702. PMID 9000754.
- Chaw, S.M.; Parkinson, C.L.; Cheng, Y.; Vincent, T.M.; Palmer, J.D. (2000). "Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers". Proceedings of the National Academy of Sciences USA. 97 (8): 4086–4091. Bibcode:2000PNAS...97.4086C. doi:10.1073/pnas.97.8.4086. PMC 18157. PMID 10760277.
- Goremykin, V.; Bobrova, V.; Pahnke, J.; Troitsky, A.; Antonov, A.; Martin, W. (1996). "Noncoding sequences from the slowly evolving chloroplast inverted repeat in addition to rbcL data do not support gnetalean affinities of angiosperms". Molecular Biology and Evolution. 13 (2): 383–396. doi:10.1093/oxfordjournals.molbev.a025597. PMID 8587503.
- Hajibabaei, M.; Xia, J.; Drouin, G. (2006). "Seed plant phylogeny: Gnetophytes are derived conifers and a sister group to Pinaceae". Molecular Phylogenetics and Evolution. 40 (1): 208–217. doi:10.1016/j.ympev.2006.03.006. PMID 16621615.
- Hansen, A.; Hansmann, S.; Samigullin, T.; Antonov, A.; Martin, W. (1999). "Gnetum and the angiosperms: molecular evidence that their shared morphological characters are convergent rather than homologous". Molecular Biology and Evolution. 16 (7): 1006–1009. doi:10.1093/oxfordjournals.molbev.a026176.
- Magallon, S.; Sanderson, M.J. (2002). "Relationships among seed plants inferred from highly conserved genes: sorting conflicting phylogenetic signals among ancient lineages". American Journal of Botany. 89 (12): 1991–2006. doi:10.3732/ajb.89.12.1991. JSTOR 4122754. PMID 21665628.
- Qiu, Y.L.; Lee, J.; Bernasconi-Quadroni, F.; Soltis, D.E.; Soltis, P.S.; Zanis, M.; Zimmer, E.A.; Chen, Z.; Savalainen, V. & Chase, M.W. (1999). "The earliest angiosperms: evidence from mitochondrial, plastid and nuclear genomes". Nature. 402 (6760): 404–407. Bibcode:1999Natur.402..404Q. doi:10.1038/46536. PMID 10586879. S2CID 4380796.
- Samigullin, T.K.; Martin, W.F.; Troitsky, A.V.; Antonov, A.S. (1999). "Molecular data from the chloroplast rpoC1 gene suggest a deep and distinct dichotomy of contemporary spermatophytes into two monophyla: gymnosperms (including Gnetalaes) and angiosperms". Journal of Molecular Evolution. 49 (3): 310–315. Bibcode:1999JMolE..49..310S. doi:10.1007/PL00006553. PMID 10473771. S2CID 4232968.
- Sanderson, M.J.; Wojciechowski, M.F.; Hu, J.M.; Sher Khan, T.; Brady, S.G. (2000). "Error, bias, and long-branch attraction in data for two chloroplast photosystem genes in seed plants". Molecular Biology and Evolution. 17 (5): 782–797. doi:10.1093/oxfordjournals.molbev.a026357. PMID 10779539.
- Rydin, C.; Kallersjo, M.; Friist, E.M. (2002). "Seed plant relationships and the systematic position of Gnetales based on nuclear and chloroplast DNA: conflicting data, rooting problems, and the monophyly of conifers". International Journal of Plant Sciences. 163 (2): 197–214. doi:10.1086/338321. JSTOR 3080238. S2CID 84578578.
- Braukmann, T.W.A.; Kuzmina, M.; Stefanovic, S. (2009). "Loss of all plastid nhd genes in Gnetales and conifers: extent and evolutionary significance for the seed plant phylogeny". Current Genetics. 55 (3): 323–337. doi:10.1007/s00294-009-0249-7. PMID 19449185. S2CID 3939394.
- Burleigh, J.G.; Mathews, S. (2007). "Phylogenetic signal in nucleotide data from seed plants: implications for resolving the seed plant tree of life". American Journal of Botany. 168 (10): 125–135. doi:10.3732/ajb.91.10.1599. PMID 21652311.
- Zhang Y, Liu Z. Genic evidence that gnetophytes are sister to all other seed plants[J]. bioRxiv, 2019: 629915. doi: https://doi.org/10.1101/629915
- Chen, Z.-D., Yang, T., Lin, L., Lu, L.-M., Li, H.-L., … Sun, M. (2016). Tree of life for the genera of Chinese vascular plants. Journal of Systematics and Evolution, 54(4), 277–306. https://doi.org/10.1111/jse.12219
- Krassilov, V.A.; Dilcher, D.L.; Douglas, J.G. (January 1998). "New ephedroid plant from the Lower Cretaceous Koonwarra Fossil Bed, Victoria, Australia". Alcheringa: An Australasian Journal of Palaeontology. 22 (2): 123–133. doi:10.1080/03115519808619195. ISSN 0311-5518.
- Yang, Yong; Wang, Yingwei; Ferguson, David Kay (2020-02-04). "A new macrofossil ephedroid plant with unusual bract morphology from the Lower Cretaceous Jiufotang Formation of northeastern China". BMC Evolutionary Biology. 20 (1): 19. doi:10.1186/s12862-019-1569-y. ISSN 1471-2148. PMC 7001366. PMID 32019502.
- Yang, Yong; Lin, Longbiao; Ferguson, David K.; Wang, Yingwei (December 2018). "Macrofossil evidence unveiling evolution of male cones in Ephedraceae (Gnetidae)". BMC Evolutionary Biology. 18 (1): 125. doi:10.1186/s12862-018-1243-9. ISSN 1471-2148. PMC 6116489. PMID 30157769.
- Yang, Yong; Ferguson, David K. (October 2015). "Macrofossil evidence unveiling evolution and ecology of early Ephedraceae". Perspectives in Plant Ecology, Evolution and Systematics. 17 (5): 331–346. doi:10.1016/j.ppees.2015.06.006.
- Yang, Yong; Lin, Longbiao; Wang, Qi (2013-03-27). "Chengia laxispicatagen. et sp. nov., a new ephedroid plant from the Early Cretaceous Yixian Formation of western Liaoning, Northeast China: evolutionary, taxonomic, and biogeographic implications". BMC Evolutionary Biology. 13 (1): 72. doi:10.1186/1471-2148-13-72. ISSN 1471-2148. PMC 3626868. PMID 23530702.
- Guo, Shuang-Xing; Sha, Jin-Geng; Bian, Li-Zeng; Qiu, Yin-Long (March 2009). "Male spike strobiles with Gnetum affinity from the Early Cretaceous in western Liaoning, Northeast China". Journal of Systematics and Evolution. 47 (2): 93–102. doi:10.1111/j.1759-6831.2009.00007.x.
- Friis, Else Marie; Pedersen, Kaj Raunsgaard; Crane, Peter R. (2014-07-03). "Welwitschioid diversity in the Early Cretaceous: evidence from fossil seeds with pollen from Portugal and eastern North America". Grana. 53 (3): 175–196. doi:10.1080/00173134.2014.915980. ISSN 0017-3134.
- Yang, Yong; Lin, Long-Biao; Ferguson, David K.; Zhang, Shou-Zhou; Wan, Tao (June 2017). "A new gnetalean macrofossil from the Early Cretaceous and its evolutionary significance". Cretaceous Research. 74: 56–64. doi:10.1016/j.cretres.2017.02.007.
- Yang, Yong; Xie, Lei; Ferguson, David K. (October 2017). "Protognetaceae: A new gnetoid macrofossil family from the Jurassic of northeastern China". Perspectives in Plant Ecology, Evolution and Systematics. 28: 67–77. doi:10.1016/j.ppees.2017.08.001.
- Ricardi-Branco, Fresia; Torres, Margarita; S., Sandra; de Souza, Ismar; E. Tavares, Paulo G.; Arruda Campos, Antonio C. (2013-05-22), Ray, Pallav (ed.), "Itajuba yansanae Gen and SP NOV of Gnetales, Araripe Basin (Albian-Aptian) in Northeast Brazil", Climate Change and Regional/Local Responses, InTech, doi:10.5772/55704, ISBN 978-953-51-1132-0, retrieved 2020-12-05
- Rothwell, Gar W.; Stockey, Ruth A. (March 2013). "Evolution and Phylogeny of Gnetophytes: Evidence from the Anatomically Preserved Seed Cone Protoephedrites eamesii gen. et sp. nov. and the Seeds of Several Bennettitalean Species". International Journal of Plant Sciences. 174 (3): 511–529. doi:10.1086/668688. ISSN 1058-5893. S2CID 84063572.
- Rydin, Catarina; Friis, Else Marie (2010-06-17). "A new Early Cretaceous relative of Gnetales: Siphonospermum simplexgen. et sp. nov. from the Yixian Formation of Northeast China". BMC Evolutionary Biology. 10 (1): 183. doi:10.1186/1471-2148-10-183. ISSN 1471-2148. PMC 2900273. PMID 20565755.
Other Sources:
- Gifford, Ernest M., Adriance S. Foster. 1989. Morphology and Evolution of Vascular Plants. Third edition. WH Freeman and Company, New York.
- Hilton, Jason, and Richard M. Bateman. 2006. Pteridosperms are the backbone of seed-plant phylogeny. Journal of the Torrey Botanical Society 133: 119-168 (abstract)