History of speciation
The scientific study of speciation — how species evolve to become new species — began around the time of Charles Darwin in the middle of the 19th century. Many naturalists at the time recognized the relationship between biogeography (the way species are distributed) and the evolution of species. The 20th century saw the growth of the field of speciation, with major contributors such as Ernst Mayr researching and documenting species' geographic patterns and relationships. The field grew in prominence with the modern evolutionary synthesis in the early part of that century. Since then, research on speciation has expanded immensely.

| Part of a series on |
| Evolutionary biology |
|---|
 |
|
The language of speciation has grown more complex. Debate over classification schemes on the mechanisms of speciation and reproductive isolation continue. The 21st century has seen a resurgence in the study of speciation, with new techniques such as molecular phylogenetics and systematics. Speciation has largely been divided into discrete modes that correspond to rates of gene flow between two incipient populations. Today however, research has driven the development of alternative schemes and the discovery of new processes of speciation.
Early history
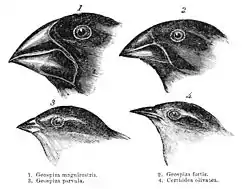
Charles Darwin introduced the idea that species could evolve and split into separate lineages, referring to it as specification in his 1859 book On the Origin of Species.[2] It was not until 1906 that the modern term speciation was coined by the biologist Orator F. Cook.[2][3] Darwin, in his 1859 publication, focused primarily on the changes that can occur within a species, and less on how species may divide into two.[4]:1 It is almost universally accepted that Darwin's book did not directly address its title.[1] Darwin instead saw speciation as occurring by species entering new ecological niches.[4]:125
Darwin's views
Controversy exists as to whether Charles Darwin recognized a true geographical-based model of speciation in his publication On the Origin of Species.[5] In chapter 11, "Geographical Distribution", Darwin discusses geographic barriers to migration, stating for example that "barriers of any kind, or obstacles to free migration, are related in a close and important manner to the differences between the productions of various regions [of the world]".[6] F. J. Sulloway contends that Darwin's position on speciation was "misleading" at the least[7] and may have later misinformed Wagner and David Starr Jordan into believing that Darwin viewed sympatric speciation as the most important mode of speciation.[4]:83 Nevertheless, Darwin never fully accepted Wagner's concept of geographical speciation.[5]
The evolutionary biologist James Mallet maintains that the mantra repeated concerning Darwin's Origin of Species book having never actually discussed speciation is specious.[1] The claim began with Thomas Henry Huxley and George Romanes (contemporaries of Darwin's), who declared that Darwin failed to explain the origins of inviability and sterility in hybrids.[1][8] Similar claims were promulgated by the mutationist school of thought during the late 20th century, and even after the modern evolutionary synthesis by Richard Goldschmidt.[1][8] Another strong proponent of this view about Darwin came from Mayr.[1][8] Mayr maintained that Darwin was unable to address the problem of speciation, as he did not define species using the biological species concept.[9] However, Mayr's view has not been entirely accepted, as Darwin's transmutation notebooks contained writings concerning the role of isolation in the splitting of species.[9] Furthermore, Many of Darwin's ideas on speciation largely match the modern theories of both adaptive radiation and ecological speciation.[5]
Biogeographic influence

Recognition of geographic factors involved in species populations was present even before Darwin, with many naturalists aware of the role of isolation in species relationships.[10]:482 In 1833, C. L. Gloger published The Variation of Birds Under the Influence of Climate in which he described geographic variations, but did not recognize that geographic isolation was an indicator of past speciation events.[10]:482 Another naturalist in 1856, Wollaston, studied island beetles in comparison to mainland species.[10]:482 He saw isolation as key to their differentiation.[10]:482 However, he did not recognize that the pattern was due to speciation.[10]:483 One naturalist, Leopold von Buch (1825) did recognize the geographic patterns and explicitly stated that geographic isolation may lead to species separating into new species.[10]:483 Mayr suggests that Von Buch was likely the first naturalist to truly suggest geographic speciation.[11] Other naturalists, such as Henry Walter Bates (1863), recognized and accepted the patterns as evidence of speciation, but in Bate's case, did not propose a coherent model.[10]:484
In 1868, Moritz Wagner was the first to propose the concept of geographic speciation[12][10]:484 in which he used the term Separationstheorie.[5] Edward Bagnall Poulton, the evolutionary biologist and a strong proponent of the importance of natural selection, highlighted the role of geographic isolation in promoting speciation,[13] in the process coining the term "sympatric speciation" in 1904.[14][15]
Wagner and other naturalists who studied the geographic distributions of animals, such as Karl Jordan and David Starr Jordan, noticed that closely related species were often geographically isolated from one another (allopatrically distributed) which lead to the advocation of the importance of geographic isolation in the origin of species.[4]:2 Karl Jordan is thought to have recognized the unification of mutation and isolation in the origin of new species — in stark contrast to the prevailing views at the time.[10]:486 David Starr Jordan reiterated Wagner's proposal in 1905, providing a wealth of evidence from nature to support the theory,[12][16][4]:2 and asserting that geographic isolation is obvious but had been unfortunately ignored by most geneticists and experimental evolutionary biologists at the time.[10]:487 Joel Asaph Allen suggested the observed pattern of geographic separation of closely related species be called "Jordan's Law" (or Wagner's Law).[10]:487 Despite the contentions, most taxonomists did accept the geographic model of speciation.[10]:487
Many of the early terms used to describe speciation were outlined by Ernst Mayr.[17] He was the first to encapsulate the then contemporary literature in his 1942 publication Systematics and the Origin of Species, from the Viewpoint of a Zoologist and in his subsequent 1963 publication Animal Species and Evolution. Like Jordan's works, they relied on direct observations of nature, documenting the occurrence of geographic speciation.[4]:86 He described the three modes: geographic, semi-geographic, and non-geographic; which today, are referred to as allopatric, parapatric, and sympatric respectively.[17] Mayr's 1942 publication, influenced heavily by the ideas of Karl Jordan and Poulton, was regarded as the authoritative review of speciation for over 20 years—and is still valuable today.[15]
A major focus of Mayr's works was on the importance of geography in facilitating speciation; with islands often acting as a central theme to many of the speciation concepts put forth.[18] One of which was the concept of peripatric speciation, a variant of allopatric speciation[19][20] (he has since distinguished the two modes by referring to them as peripatric and dichopatric[21]). This concept arose by an interpretation of Wagner's Separationstheorie as a form of founder effect speciation that focused on small geographically isolated species.[5] This model was later expanded and modified to incorporate sexual selection by Kenneth Y. Kaneshiro in 1976 and 1980.[22][23][24]
Modern evolutionary synthesis
Many geneticists at the time did little to bridge the gap between the genetics of natural selection and the origin of reproductive barriers between species.[4]:3 Ronald Fisher proposed a model of speciation in his 1930 publication The Genetical Theory of Natural Selection, where he described disruptive selection acting on sympatric or parapatric populations — with reproductive isolation completed by reinforcement.[25] Other geneticists such as J. B. S. Haldane did not even recognize that species were real, while Sewall Wright ignored the topic, despite accepting allopatric speciation.[4]:3
The primary contributors to the incorporation of speciation into modern evolutionary synthesis were Ernst Mayr and Theodosius Dobzhansky.[25] Dobzhansky, a geneticist, published Genetics and the Origin of Species in 1937, in which he formulated the genetic framework for how speciation could occur.[4]:2 He recognized that speciation was an unsolved problem in biology at the time, rejecting Darwin's position that new species arose by occupation of new niches — contending that reproductive isolation was instead based on barriers to gene flow.[4]:2 Subsequently, Mayr conducted extensive work on the geography of species, emphasizing the importance of geographic separation and isolation, in which he filled Dobzhansky's gaps concerning the origin of biodiversity (in his 1942 book).[26] Both of their works gave rise, not without controversy, to the modern understanding of speciation; stimulating a wealth of research on the topic.[4]:3 Furthermore, this extended to plants as well as animals with G. Ledyard Stebbins’s book, Variation and Evolution in Plants and the much later, 1981 book, Plant Speciation by Verne Grant.

In 1947, "a consensus had been achieved among geneticists, paleontologists and systematists and that evolutionary biology as an independent biological discipline had been established" during a Princeton University conference.[28] This 20th century synthesis incorporated speciation. Since then, the ideas have been consistently and repeatedly confirmed.[26]
Contemporary work
After the synthesis, speciation research continued largely within natural history and biogeography — with much less emphasis on genetics.[4]:4 The study of speciation has seen its largest increase since the 1980s[4]:4 with an influx of publications and a host of new terms, methods, concepts, and theories.[17] This "third phase" of work — as Jerry A. Coyne and H. Allen Orr put it — has led to a growing complexity of the language used to describe the many processes of speciation.[17] The research and literature on speciation have become, "enormous, scattered, and increasingly technical".[4]:1
From the 1980s, new research tools increased the robustness of research,[4]:4 assisted by new methods, theoretical frameworks, models, and approaches.[17] Coyne and Orr discuss the modern, post-1980s developments centered around five major themes:
- genetics (also a primary factor in the Modern Synthesis),
- molecular biology and analysis (namely, phylogenetics and systematics);
- comparative analysis;
- mathematical modeling and computer simulations; and
- the role of ecology.[4]:5
Ecologists became aware that the ecological factors behind speciation were under-represented. This saw the growth in research concerning ecology's role in facilitating speciation — rightly designated ecological speciation.[4]:4 This focus on ecology generated a host of new terms relating to the barriers to reproduction[17] (e.g. allochronic speciation, in which gene flow is reduced or removed by timing of breeding periods; or habitat isolation, in which species occupy different habitats within the same area). Sympatric speciation, regarded by Mayr as unlikely, has become widely accepted.[29][30][31] Research on the influence of natural selection on speciation, including the process of reinforcement, has grown.[32]
Researchers have long debated the roles of sexual selection, natural selection, and genetic drift in speciation.[4]:383 Darwin extensively discussed sexual selection, with his work greatly expanded on by Ronald Fisher; however, it was not until 1983 that the biologist Mary Jane West-Eberhard recognized the importance of sexual selection in speciation.[33][4]:3 Natural selection plays a role in that any selection towards reproductive isolation can result in speciation — whether indirectly or directly. Genetic drift has been widely researched from the 1950s onwards, especially with peak-shift models of speciation by genetic drift.[4]:388 Mayr championed founder effects, in which isolated individuals, like those found on islands near a mainland, experience a strong population bottleneck, as they contain only a small sample of the genetic variation in the main population.[4]:390[34] Later, other biologists such as Hampton L. Carson, Alan Templeton, Sergey Gavrilets, and Alan Hastings developed related models of speciation by genetic drift, noting that islands were inhabited mostly by endemic species.[35] Selection's role in speciation is widely supported, whereas founder effect speciation is not,[4]:410 having been subject to a number of criticisms.[36]
Classification debate
.png.webp)

Throughout the history of research concerning speciation, classification and delineation of modes and processes have been debated. Julian Huxley divided speciation into three separate modes: geographical speciation, genetic speciation, and ecological speciation.[10]:427 Sewall Wright proposed ten different, varying modes.[10]:427 Ernst Mayr championed the importance of physical, geographic separation of species populations, maintaining it to be of major importance to speciation. He originally proposed the three primary modes known today: geographic, semi-geographic, non-geographic;[17] corresponding to allopatric, parapatric, and sympatric respectively.
The phrase "modes of speciation" is imprecisely defined, most often indicating speciation occurring as a result of a species geographic distribution.[37] More succinctly, the modern classification of speciation is often described as occurring on a gene flow continuum (i.e., allopatry at and sympatry at [38][39]) This gene flow concept views speciation as based on the exchange of genes between populations instead of seeing a purely geographic setting as necessarily relevant. Despite this, concepts of biogeographic modes can be translated into models of gene flow (such as that in the image at left); however, this translation has led to some confusion of language in the scientific literature.[17]


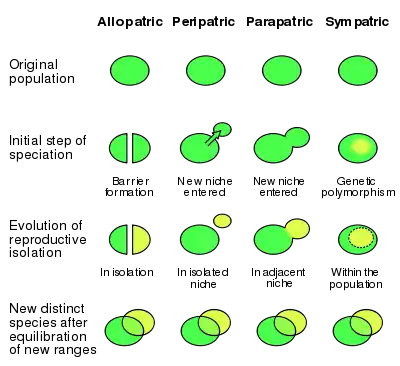
As research has expanded over the decades, the geographic scheme has been challenged. The traditional classification is considered by some researchers to be obsolete,[40] while others argue for its merits. Proponents of non-geographic schemes often justify non-geographic classifications, not by rejection of the importance of reproductive isolation (or even the processes themselves), but instead by the fact that it simplifies the complexity of speciation.[41] One major critique of the geographic framework is that it arbitrarily separates a biological continuum into discontinuous groups.[41] Another criticism rests with the fact that, when speciation is viewed as a continuum of gene flow, parapatric speciation becomes unreasonably represented by the entire continuum[42]—with allopatric and sympatric existing in the extremes.[41] Coyne and Orr argue that the geographic classification scheme is valuable in that biogeography controls the strength of the evolutionary forces at play, as gene flow and geography are clearly linked.[40] James Mallet and colleagues contend that the sympatric vs. allopatric dichotomy is valuable to determine the degree in which natural selection acts on speciation.[43] Kirkpatrick and Ravigné categorize speciation in terms of its genetic basis or by the forces driving reproductive isolation.[4]:85 Here, the geographic modes of speciation are classified as types of assortive mating.[44] Fitzpatrick and colleagues believe that the biogeographic scheme "is a distraction that could be positively misleading if the real goal is to understand the influence of natural selection on divergence."[40] They maintain that, to fully understand speciation, "the spatial, ecological, and genetic factors" involved in divergence must be explored.[40] Sara Via recognizes the importance of geography in speciation but suggests that classification under this scheme be abandoned.[30]
History of modes and mechanisms
Sympatric speciation
Sympatric speciation, from its beginnings with Darwin (who did not coin the term), has been a contentious issue.[37][4]:125 Mayr, along with many other evolutionary biologists, interpreted Darwins's view of speciation and the origin of biodiversity as arising by species entering new ecological niches—a form of sympatric speciation.[1] Before Mayr, sympatric speciation was regarded as the primary mode of speciation. In 1963, Mayr provided a strong criticism, citing various flaws in the theory.[4]:126 After that, sympatric speciation fell out of favor with biologists and has only recently seen a resurgence in interest.[4]:126 Some biologists, such as James Mallet, believe that Darwin's view on speciation was misunderstood and misconstrued by Mayr.[1][45] Today, sympatric speciation is supported by evidence from laboratory experiments and observations from nature.[4]:127[29]
Hybrid speciation
For most of the history of speciation, hybridization (polyploidy) has been a contentious issue, as botanists and zoologists have traditionally viewed hybridization's role in speciation differently.[17] Carl Linnaeus was the earliest to suggest hybridization in 1760,[46] Øjvind Winge was the first to confirm allopolyploidy in 1917,[46][47] and a later experiment conducted by Clausen and Goodspeed in 1925 confirmed the findings.[46] Today it is widely recognized as a common mechanism of speciation.[48]
Historically, zoologists considered hybridization to be a rare phenomenon, while botanists found it to be commonplace in plant species.[17] The botanists G. Ledyard Stebbins and Verne Grant were two of the well known botanists who championed the idea of hybrid speciation during the 1950s to the 1980s.[17] Hybrid speciation, also called polyploid speciation (or polyploidy) is speciation that results by an increase in the number of sets of chromosomes.[4]:321 It is effectively a form of sympatric speciation that happens instantly.[4]:322 Grant coined the term recombinational speciation in 1981; a special form of hybrid speciation where a new species results from hybridization and is itself, reproductively isolated from both its parents.[4]:337 Recently, biologists have increasingly recognized that hybrid speciation can occur in animals as well.[49]
Reinforcement

The concept of speciation by reinforcement has a complex history, with its popularity among scholars changing significantly over time.[32][4]:353 The theory of reinforcement experienced three phases of historical development:[4]:366
- plausibility based on unfit hybrids
- implausibility based on the finding that hybrids may have some fitness
- plausibility based on empirical studies and biologically complex and realistic models
It was originally proposed by Alfred Russel Wallace in 1889,[4]:353 termed the Wallace effect—a term rarely used by scientists today.[50] Wallace's hypothesis differed from the modern conception in that it focused on post-zygotic isolation, strengthened by group selection.[4]:353[51][52] Dobzhansky was the first to provide a thorough, modern description of the process in 1937,[4]:353 though the actual term itself was not coined until 1955 by W. Frank Blair.[53]
In 1930, Ronald Fisher laid out the first genetic description of the process of reinforcement in The Genetical Theory of Natural Selection, and in 1965 and 1970 the first computer simulations were run to test for its plausibility.[4]:366 Later, population genetic[54] and quantitative genetic[55] studies were conducted showing that completely unfit hybrids lead to an increase in pre-zygotic isolation.[4]:368 After Dobzhansky's idea rose to the forefront of speciation research, it garnered significant support—with Dobzhansky suggesting that it illustrated the final step in speciation (e.g. after an allopatric population comes into secondary contact).[4]:353 In the 1980s, many evolutionary biologists began to doubt the plausibility of the idea,[4]:353 based not on empirical evidence, but largely on the growth of theory that deemed it an unlikely mechanism of reproductive isolation.[56] A number of theoretical objections arose at the time. Since the early 1990s, reinforcement has seen a revival in popularity, with perceptions by evolutionary biologists accepting its plausibility—due primarily from a sudden increase in data, empirical evidence from laboratory studies and nature, complex computer simulations, and theoretical work.[4]:372–375
The scientific language concerning reinforcement has also differed over time, with different researchers applying various definitions to the term.[50] First used to describe the observed mating call differences in Gastrophryne frogs within a secondary contact hybrid zone,[50] reinforcement has also been used to describe geographically separated populations that experience secondary contact.[57] Roger Butlin demarcated incomplete post-zygotic isolation from complete isolation, referring to incomplete isolation as reinforcement and completely isolated populations as experiencing reproductive character displacement.[58] Daniel J. Howard considered reproductive character displacement to represent either assortive mating or the divergence of traits for mate recognition (specifically between sympatric populations).[50] Under this definition, it includes pre-zygotic divergence and complete post-zygotic isolation.[59] Maria R. Servedio and Mohamed Noor consider any detected increase in pre-zygotic isolation as reinforcement, as long as it is a response to selection against mating between two different species.[60] Coyne and Orr contend that, "true reinforcement is restricted to cases in which isolation is enhanced between taxa that can still exchange genes".[4]:354
See also
References
- James Mallet (2008), "A century of evolution: Ernst Mayr (1904-2005): Mayr's view of Darwin: was Darwin wrong about speciation?", Biological Journal of the Linnean Society, 95 (1): 3–16, doi:10.1111/j.1095-8312.2008.01089.x
- B. N. Singh (2012), "Concepts of species and modes of speciation", Current Science, 103 (7): 784–790
- Cook, Orator F. (March 30, 1906). "Factors of species-formation". Science. 23 (587): 506–507. Bibcode:1906Sci....23..506C. doi:10.1126/science.23.587.506. ISSN 0036-8075. PMID 17789700.
- Jerry A. Coyne; H. Allen Orr (2004), Speciation, Sinauer Associates, pp. 1–545, ISBN 978-0-87893-091-3
- James Mallet (2010), "Why was Darwin's view of species rejected by twentieth century biologists?", Biology & Philosophy, 25 (4): 497–527, doi:10.1007/s10539-010-9213-7, S2CID 38621736
- Darwin, Charles (1859). On the Origin of Species. Murray. p. 347. Archived from the original on 2008-10-05.
- F. J. Sulloway (1979), "Geographic isolation in Darwin's thinking: the vicissitudes of a crucial idea", Studies in the History of Biology, 3: 23–65, PMID 11610987
- Mallet, James (2013). Darwin and species. In Michael Ruse (eds) The Cambridge Encyclopedia of Darwin and Evolutionary Thought, Cambridge University Press, Pp. 109–115.
- Malcolm J. Kottler (1978), "Charles Darwin's biological species concept and theory of geographic speciation: the transmutation notebooks", Annals of Science, 35 (3): 275–297, doi:10.1080/00033797800200251
- Ernst Mayr (1963), Animal Species and Evolution, Harvard University Press, pp. 1–797
- Ernst Mayr (1998), The Evolutionary Synthesis: Perspectives on the Unification of Biology, Harvard University Press, p. 36, ISBN 978-0674272262
- David Starr Jordan (1905), "The Origin of Species Through Isolation", Science, 22 (566): 545–562, Bibcode:1905Sci....22..545S, doi:10.1126/science.22.566.545, PMID 17832412
- Hannes Schuler, Glen R. Hood, Scott P. Egan, and Jeffrey L. Feder (2016), Meyers, Robert A (ed.), "Modes and Mechanisms of Speciation", Reviews in Cell Biology and Molecular Medicine, 2 (3): 60–93, doi:10.1002/3527600906, ISBN 9783527600908CS1 maint: multiple names: authors list (link)
- Ernst Mayr (1942), Systematics and origin of species, Columbia University Press, p. 148
- James Mallet (2004), "Perspectives: Poulton, Wallace and Jordan: how discoveries in Papilio butterflies led to a new species concept 100 years ago", Systematics and Biodiversity, 1 (4): 441–452, doi:10.1017/S1477200003001300, S2CID 86041887
- David Starr Jordan (1908), "The Law of Geminate Species", American Naturalist, 42 (494): 73–80, doi:10.1086/278905
- Richard G. Harrison (2012), "The Language of Speciation", Evolution, 66 (12): 3643–3657, doi:10.1111/j.1558-5646.2012.01785.x, PMID 23206125, S2CID 31893065
- Brent C. Emerson (2008), "A century of evolution: Ernst Mayr (1904–2005): Speciation on islands: what are we learning?", Biological Journal of the Linnean Society, 95 (1): 47–52, doi:10.1111/j.1095-8312.2008.01120.x
- Mayr, E. 1954. Change of genetic environment and evolution. In: Evolution as a Process (J. Huxley, A. C. Hardy & E. B. Ford, eds), pp. 157–180. Unwin Brothers, London.
- Mayr, E. 1982. Processes of speciation in animals. In: Mechanisms of Speciation (A. R. I. Liss, ed.), pp. 1–19. Alan R. Liss Inc., New York.
- Ernst Mayr (2001), What Evolution Is, Basic Books, pp. 178–179, ISBN 978-0465044269
- Kenneth Y. Kaneshiro (1976), "Ethological isolation and phylogeny in the Plantibia subgroup of Hawaiian Drosophila", Evolution, 30 (4): 740–745, doi:10.1111/j.1558-5646.1976.tb00954.x, PMID 28563322, S2CID 205773169
- Kenneth Y. Kaneshiro (1980), "Sexual selection, speciation and the direction of evolution", Evolution, 34 (3): 437–444, doi:10.1111/j.1558-5646.1980.tb04833.x, PMID 28568697
- Anders Ödeen and Ann-Britt Florin (2002), "Sexual selection and peripatric speciation: the Kaneshiro model revisited", Journal of Evolutionary Biology, 15 (2): 301–306, doi:10.1046/j.1420-9101.2002.00378.x, S2CID 82095639
- Jerry A. Coyne (1994), "Ernst Mayr and the origin of species", Evolution, 48 (1): 19–30, doi:10.1111/j.1558-5646.1994.tb01290.x, PMID 28567778
- Ernst Mayr (2004), "80 years of watching the evolutionary scenery" (PDF), Science, 305 (5680): 46–47, doi:10.1126/science.1100561, PMID 15232092, S2CID 161868412, archived (PDF) from the original on 2018-02-03
- Michael F. Clairidge and Vaughan Southgate (2008), "A century of evolution: Ernst Mayr (1904–2005): Introduction", Biological Journal of the Linnean Society, 95 (1): 1–2, doi:10.1111/j.1095-8312.2008.01119.x
- Jürgen Haffer (2007), Ornithology, Evolution, and Philosophy: The Life and Science of Ernst Mayr 1904–2005, Springer-Verlag Berlin Heidelberg, pp. 183–241, ISBN 978-3-540-71777-5
- Michael Turelli; Nicholas H. Barton; Jerry A. Coyne (2001), "Theory and speciation", Trends in Ecology & Evolution, 16 (7): 330–343, doi:10.1016/S0169-5347(01)02177-2, PMID 11403865
- Sara Via (2001), "Sympatric speciation in animals: the ugly duckling grows up", Trends in Ecology & Evolution, 16 (1): 381–390, doi:10.1016/S0169-5347(01)02188-7, PMID 11403871
- James Mallet (2001), "The Speciation Revolution", Journal of Evolutionary Biology, 14 (6): 887–888, doi:10.1046/j.1420-9101.2001.00342.x
- Mohamed A. F. Noor (1999), "Reinforcement and other consequences of sympatry", Heredity, 83 (5): 503–508, doi:10.1038/sj.hdy.6886320, PMID 10620021
- Marlene Zuk (2004), "2003 Sewall Wright Award: Mary Jane West‐Eberhard", American Naturalist, 163 (1): i–ii, doi:10.1086/381946
- Mayr, Ernst (1954). Change of genetic environment and evolution. In J. Huxley, A. C. Hardy, and E. B. Ford. (eds) Evolution as a Process, George Allen and Unwin, London, Pp. 157–180.
- Provine, William Ball (1989). Founder effects and genetic revolutions in microevolution and speciation. In L. V. Giddings, K. Y. Kaneshiro, and W. W. Anderson. (eds) Genetics, Speciation, and the Founder Principle, Oxford University Press, New York, Pp. 43–76.
- Matute, D. R. (2013), "The role of founder effects on the evolution of reproductive isolation", Journal of Evolutionary Biology, 26 (11): 2299–2311, doi:10.1111/jeb.12246, PMID 24118666, S2CID 10192721
- Shaw, Kerry L. (2012). "Species and Speciation: Overview". eLS. doi:10.1002/9780470015902.a0001742.pub2. ISBN 978-0470016176.
- Sergey Gavrilets (2004), Fitness landscapes and the origin of species, Princeton University Press, p. 13
- Sergey Gavrilets (2003), "Perspectiver: Models of Speciation: What have we Learned in 40 Years?", Evolution, 57 (10): 2197–2215, doi:10.1111/j.0014-3820.2003.tb00233.x, PMID 14628909, S2CID 2936776
- Benjamin M. Fitzpatrick, James A. Fordyce, and Sergey Gavrilets (2009), "Pattern, process, and geographic modes of speciation", Journal of Evolutionary Biology, 22 (11): 2342–2347, doi:10.1111/j.1420-9101.2009.01833.x, PMID 19732257, S2CID 941124CS1 maint: multiple names: authors list (link)
- Roger K. Butlin, Juan Galindo, and John W. Grahame (2008), "Sympatric, parapatric or allopatric: the most important way to classify speciation?", Philosophical Transactions of the Royal Society B: Biological Sciences, 363 (1506): 2997–3007, doi:10.1098/rstb.2008.0076, PMC 2607313, PMID 18522915CS1 maint: multiple names: authors list (link)
- Benjamin M. Fitzpatrick, James A. Fordyce, and Sergey Gavrilets (2009), "What, if anything, is sympatric speciation?", Journal of Evolutionary Biology, 21 (6): 1452–1459, doi:10.1111/j.1420-9101.2008.01611.x, PMID 18823452CS1 maint: multiple names: authors list (link)
- James Mallet, Axel Meyer, Patrik Nosil, Jeffrey L. Feder (2009), "Space, sympatry and speciation", Journal of Evolutionary Biology, 22 (11): 2332–2341, doi:10.1111/j.1420-9101.2009.01816.x, PMID 19732264CS1 maint: multiple names: authors list (link)
- Mark Kirkpatrick; Virginie Ravigné (2002), "Speciation by Natural and Sexual Selection: Models and Experiments", The American Naturalist, 159: S22–S35, doi:10.1086/338370, PMID 18707367
- James Mallet (2005), "Speciation in the 21st century", Heredity, 95 (1): 105–109, doi:10.1038/sj.hdy.6800686, ISSN 0018-067X
- Abbott, Richard J.; Reiseberg, Loren H. (2012). "Hybrid Speciation". eLS. doi:10.1002/9780470015902.a0001753.pub2. ISBN 978-0470016176.
- R. E. Clausen and T. H. Goodspeed (1925), "Interspecific Hybridization in Nicotiana. II. a Tetraploid GLUTINOSA-TABACUM Hybrid, an Experimental Verification of Winge's Hypothesis", Genetics, 10 (3): 278–284, PMC 1200860, PMID 17246274
- Douglas E. Soltis, Richard J.A. Buggs, Jeff J. Doyle, and Pamela S. Soltis (2010), "What we still don't know about polyploidy", Taxon, 59 (5): 1387–1403, doi:10.1002/tax.595006CS1 maint: multiple names: authors list (link)
- James Mallet (2007), "Hybrid speciation", Nature, 446 (7133): 279–283, Bibcode:2007Natur.446..279M, doi:10.1038/nature05706, PMID 17361174, S2CID 1016526
- Sætre, Glenn-Peter (2012). "Reinforcement". eLS. doi:10.1002/9780470015902.a0001754.pub3. ISBN 978-0470016176. Missing or empty
|title=(help) - Littlejohn, M. J. (1981). Reproductive isolation: A critical review. In W. R. Atchley and D. S. Woodruff (eds) Evolution and Speciation, Cambridge University Press, Pp. 298–334.
- Mario A. Fares (2015), Natural Selection: Methods and Applications, CRC Press, p. 3, ISBN 9781482263725
- Blair, W. Frank (1955), "Mating call and stage of speciation in the Microhyla olivacea-M. carolinensis complex", Evolution, 9 (4): 469–480, doi:10.1111/j.1558-5646.1955.tb01556.x, S2CID 88238743
- Stanley Sawyer and Daniel Hartl (1981), "On the evolution of behavioral reproductive isolation: The Wallace effect", Theoretical Population Biology, 19 (1): 261–273, doi:10.1016/0040-5809(81)90021-6
- J. A. Sved (1981), "A Two-Sex Polygenic Model for the Evolution of Premating Isolation. I. Deterministic Theory for Natural Populations", Genetics, 97 (1): 197–215, PMC 1214384, PMID 17249073
- Jeremy L. Marshall, Michael L. Arnold, and Daniel J. Howard (2002), "Reinforcement: the road not taken", Trends in Ecology & Evolution, 17 (12): 558–563, doi:10.1016/S0169-5347(02)02636-8CS1 maint: multiple names: authors list (link)
- Theodosius Dobzhansky (1937), Genetics And the Origin of Species, Columbia University Press
- Butlin, Roger K. (1989). Reinforcement of premating isolation. In Otte, D. and Endler, John A. (eds) Speciation and its Consequences, Sinauer Associates, pp. 158–179, ISBN 0-87893-657-2
- Howard, D. J. (1993). Reinforcement: origin, dynamics and fate of an evolutionary hypothesis. In: Harrison, R. G. (eds) Hybrid Zones and the Evolutionary Process, Oxford University Press, pp. 46–69.
- Maria R. Servedio and Mohamed A. F. Noor (2003), "The Role of Reinforcement in Speciation: Theory and Data", Annual Review of Ecology, Evolution, and Systematics, 34: 339–364, doi:10.1146/annurev.ecolsys.34.011802.132412
