Myriapoda
Myriapoda is a subphylum of arthropods containing millipedes, centipedes, and others. The group contains over 16,000 species, most of which are terrestrial.[2] Although their name suggests they have myriad (10,000) legs, myriapods range from having up to 750 legs (the millipede Illacme plenipes)[3] to having fewer than ten legs.
| Myriapods | |
|---|---|
 | |
| Representatives of the four extant myriapod classes. Clockwise from top left: Chilopoda, Diplopoda, Symphyla, and Pauropoda. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Subphylum: | Myriapoda Latreille, 1802 |
| Classes[1] | |
The fossil record of myriapods reaches back into the late Silurian, although molecular evidence suggests a diversification in the Cambrian Period,[4] and Cambrian fossils exist which resemble myriapods.[2] The oldest unequivocal myriapod fossil is of the millipede Pneumodesmus newmani, from the late Silurian (428 million years ago). P. newmani is also important as the earliest known terrestrial animal.[5][6] The phylogenetic classification of myriapods is still debated.
The scientific study of myriapods is myriapodology, and those who study myriapods are myriapodologists.[7]
Anatomy

Myriapods have a single pair of antennae and, in most cases, simple eyes. Exceptions include the large and well-developed compound eyes of Scutigera [8] The mouthparts lie on the underside of the head, with an "epistome" and labrum forming the upper lip, and a pair of maxillae forming the lower lip. A pair of mandibles lie inside the mouth. Myriapods breathe through spiracles that connect to a tracheal system similar to that of insects. There is a long tubular heart that extends through much of the body, but usually few, if any, blood vessels.[9]
Malpighian tubules excrete nitrogenous waste into the digestive system, which typically consists of a simple tube. Although the ventral nerve cord has a ganglion in each segment, the brain is relatively poorly developed.[9]
During mating, male myriapods produce a packet of sperm, or spermatophore, which they must transfer to the female externally; this process is often complex and highly developed. The female lays eggs which hatch as much-shortened versions of the adults, with only a few segments and as few as three pairs of legs. With the exception of the two centipede orders Scolopendromorpha and Geophilomorpha, which have epimorphic development (all body segments are formed segments embryonically), the young add additional segments and limbs as they repeatedly moult to reach the adult form.[9] The process of adding new segments during postembryonic growth is known as anamorphosis, of which there are three types; euanamorphosis, where every moult is followed by addition of new segments, even after reaching sexual maturity. Emianamorphosis, where new segments are added until a certain stadium, and further moults happens without addition of segments. And teloanamorphosis, where the addition of new segments stops after the adult form is reached, after which no further moults occur.[10]
Ecology
Myriapods are most abundant in moist forests, where they fulfill an important role in breaking down decaying plant material,[2] although a few live in grasslands, semi-arid habitats or even deserts.[11] A very small percentage of species are littoral (found along the sea shore).[12][13] The majority are detritivorous, with the exception of centipedes, which are chiefly nocturnal predators. A few species of centipedes and millipedes are able to produce light and are therefore bioluminescent [14] Pauropodans and symphylans are small, sometimes microscopic animals that resemble centipedes superficially and live in soils. Millipedes differ from the other groups in having their body segments fused into pairs, giving the appearance that each segment bears two pairs of legs, while the other three groups have a single pair of legs on each body segment.
Although not generally considered dangerous to humans, many millipedes produce noxious secretions (often containing benzoquinones) which in rare cases can cause temporary blistering and discolouration of the skin.[15] Large centipedes, however, can bite humans, and although the bite may cause intense pain and discomfort, fatalities are extremely rare.[16]
Classification
There has been much debate as to which arthropod group is most closely related to the Myriapoda.[17] Under the Mandibulata hypothesis, Myriapoda is the sister taxon to Pancrustacea, a group comprising the Crustacea and Hexapoda (insects and their close relatives). Under the Atelocerata hypothesis, Hexapoda is the closest, whereas under the Paradoxopoda hypothesis, Chelicerata is the closest. This last hypothesis, although supported by few, if any, morphological characters, is supported by a number of molecular studies.[18] A 2020 study found numerous characters of the eye and preoral region suggesting that the closest relatives to crown myriapods are the extinct Euthycarcinoids.[19] There are four classes of extant myriapods, Chilopoda (centipedes), Diplopoda, Pauropoda and Symphyla, containing a total of around 12,000 species.[20] While each of these groups of myriapods is believed to be monophyletic, relationships among them are less certain.[21]
Centipedes

Centipedes make up the class Chilopoda. They are fast, predatory and venomous, hunting mostly at night. There are around 3,300 species,[20] ranging from the diminutive Nannarrup hoffmani (less than 12 mm or 1⁄2 in in length)[22] to the giant Scolopendra gigantea, which may exceed 30 centimetres (12 in).
Millipedes

Millipedes form the class Diplopoda. Most millipedes are slower than centipedes, and feed on leaf litter and detritus. They are distinguished by the fusion of each pair of body segments into a single unit, giving the appearance of having two pairs of legs per segment. Around 12,000 species have been described, which may represent less than a tenth of the true global millipede diversity.[20] The name "millipede" is a compound word formed from the Latin roots millia ("thousand") and pes (gen. pedis) ("foot"), although millipedes typically have between 36 and 400 legs. One species, Illacme plenipes, has the greatest number of legs of any animal, with 750.[3] Pill millipedes are much shorter, and are capable of rolling up into a ball, like pillbugs.
Symphyla
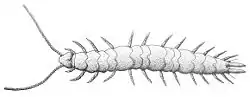
About 200 species of them are known worldwide.[20] They resemble centipedes but are smaller and translucent. Many spend their lives as soil infauna, but some live arboreally. Juveniles have six pairs of legs, but, over a lifetime of several years, add an additional pair at each moult so that the adult instar has twelve pairs of legs.[23]
Pauropoda
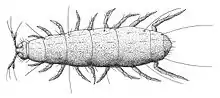
Pauropoda is another small group of small myriapods. They are typically 0.5–2.0 mm long and live in the soil on all continents except Antarctica.[24] Over 700 species have been described.[20] They are believed to be the sister group to millipedes, and have the dorsal tergites fused across pairs of segments, similar to the more complete fusion of segments seen in millipedes.[25]
Arthropleuridea
Arthropleurideans were ancient myriapods that are now extinct, known from the late Silurian to the Permian. The most famous members are from the genus Arthropleura, which was a giant, probably herbivorous, animal that could be up to 3 metres (10 ft) long, but the group also includes species less than 1 cm (0.39 in). Arthropleuridea was historically considered a distinct class of myriapods, but since 2000 scientific consensus has viewed the group as a subset of millipedes, although the relationship of arthropleurideans to other millipedes and to each other is debated.[26][27]
Myriapod relationships

A variety of groupings (clades) of the myriapod classes have been proposed, and married, some of which are mutually exclusive, and all of which represent hypotheses of evolutionary relationships. Traditional relationships supported by morphological similarities (anatomical or developmental similarities) are challenged by newer relationships supported by molecular evidence (including DNA sequence and amino acid similarities).[28][29]
- Dignatha (also called Collifera) is a clade consisting of millipedes and pauropods, and is supported by morphological similarities including the presence of a gnathochilarium (a modified jaw and plate apparatus) and a collum, a legless segment behind the head.
- Trignatha (also called Atelopoda) is a grouping of centipedes and symphylans, united by similarities of mouthparts.
- Edafopoda is a grouping of symphylans and pauropodans that is supported by shared genetic sequences, yet conflicts with Dignatha and Trignatha.[30]
- Progoneata is a group encompassing millipedes, pauropods and symphylans while excluding centipedes. Shared features include reproductive openings (gonopores) behind the second body segment, and sensory hairs (trichobothria) with a bulb-like swelling. It is compatible with either Dignatha or Edafopoda.[29]
See also
- Euthycarcinoidea, a group of enigmatic arthropods that may be ancestral to myriapods
- Colonization of land, major evolutionary stages leading to terrestrial organisms
- Metamerism, the condition of multiple linearly repeated body segments
References
- "Myriapoda". Integrated Taxonomic Information System.
- Ben Waggoner (February 21, 1996). "Introduction to the Myriapoda". University of California, Berkeley.
- Paul E. Marek & Jason E. Bond (June 8, 2006). "Biodiversity hotspots: rediscovery of the world's leggiest animal". Nature. 441 (7094): 707. Bibcode:2006Natur.441..707M. doi:10.1038/441707a. PMID 16760967. S2CID 4398951.
- Markus Friedrich & Diethard Tautz (2002). "Ribosomal DNA phylogeny of the major extant arthropod classes and the evolution of myriapods". Nature. 376 (6536): 165–167. Bibcode:1995Natur.376..165F. doi:10.1038/376165a0. PMID 7603566. S2CID 4270910.
- Rowland Shelley & Paul Marek (March 1, 2005). "Millipede Fossils". East Carolina University. Archived from the original on May 27, 2011.
- Garwood, Russell J.; Edgecombe, Gregory D. (September 2011). "Early Terrestrial Animals, Evolution, and Uncertainty". Evolution: Education and Outreach. 4 (3): 489–501. doi:10.1007/s12052-011-0357-y.
- Sue Hubbell (2000). Waiting for Aphrodite: Journeys Into the Time Before Bones. Houghton Mifflin Harcourt. p. 53. ISBN 978-0-618-05684-2.
- Müller, CHG; Rosenberg, J; Richter, S; Meyer-Rochow, VB (2003). "The compound eye of Scutigera coleoptrata (Linnaeus, 1758) (Chilopoda; Notostigmophora): an ultrastructural re-investigation that adds support to the Mandibulata concept". Zoomorphology. 122 (4): 191–209. doi:10.1007/s00435-003-0085-0.
- Robert D. Barnes (1982). Invertebrate Zoology. Philadelphia, PA: Holt-Saunders International. pp. 810–827. ISBN 978-0-03-056747-6.
- Anamorphosis in millipedes (Diplopoda)—the present state of knowledge with some developmental and phylogenetic considerations
- "Myriapod". Britannica Concise Encyclopedia.
- Barber, A.D. (2009). "Littoral myriapods: a review" (PDF). Soil Organisms. 81 (3): 735–760. Archived from the original (PDF) on 2016-03-03. Retrieved 2013-11-05.
- Barber, A.D. (Ed) (2013). "World Database of Littoral Myriapoda". World Register of Marine Species. Retrieved 25 October 2013.CS1 maint: extra text: authors list (link)
- Rosenberg, Joerg; Meyer-Rochow, Victor Benno (2009). Meyer-Rochow V.B. (ed.). Bioluminescence in Focus - a collection of illuminating essays. Research Signpost; Trivandrum, Kerala, India. pp. 139–147.
- "Strange and Unusual Millipedes". herper.com. Archived from the original on April 2, 2012. Retrieved July 2, 2007.
- Sean P. Bush; Bradley O. King; Robert L. Norris; Scott A. Stockwell (2001). "Centipede envenomation". Wilderness & Environmental Medicine. 12 (2): 93–99. doi:10.1580/1080-6032(2001)012[0093:CE]2.0.CO;2. PMID 11434497.
- Gregory D. Edgecombe (2004). "Morphological data, extant Myriapoda, and the myriapod stem-group". Contributions to Zoology. 73 (3): 207–252. doi:10.1163/18759866-07303002.
- Alexandre Hassanin (2006). "Phylogeny of Arthropoda inferred from mitochondrial sequences: strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution". Molecular Phylogenetics and Evolution. 38 (1): 100–116. doi:10.1016/j.ympev.2005.09.012. PMID 16290034.
- Edgecombe, Gregory D.; Strullu-Derrien, Christine; Góral, Tomasz; Hetherington, Alexander J.; Thompson, Christine; Koch, Markus (2020-04-01). "Aquatic stem group myriapods close a gap between molecular divergence dates and the terrestrial fossil record". Proceedings of the National Academy of Sciences. 117 (16): 8966–8972. doi:10.1073/pnas.1920733117. ISSN 0027-8424. PMC 7183169. PMID 32253305.
- A. D. Chapman (2005). Numbers of Living Species in Australia and the World (PDF). Department of the Environment and Heritage. p. 23. ISBN 978-0-642-56850-2. Archived from the original (PDF) on 25 October 2007. Retrieved 1 January 2011.
- Jerome C. Regier, Heather M. Wilson & Jeffrey W. Shultz (2005). "Phylogenetic analysis of Myriapoda using three nuclear protein-coding genes". Molecular Phylogenetics and Evolution. 34 (1): 147–158. doi:10.1016/j.ympev.2004.09.005. PMID 15579388.
- "Central Park survey finds new centipede". American Museum of Natural History. January 29, 2003.
- "Garden Symphylans". Integrated Pest Management on Peppermint-IPMP3.0. Oregon State University. Archived from the original on August 3, 2007. Retrieved July 2, 2007.
- "Pauropods: Pauropoda". Insects and Spiders Scientific Reference. Retrieved July 2, 2007.
- David Kendall (June 6, 2005). "Pauropods & Symphylids". Kendall Bioresearch.
- Wilson, Heather M.; Shear, William A. (2000). "Microdecemplicida, a new order of minute arthropleurideans (Arthropoda: Myriapoda) from the Devonian of New York State, U.S.A.". Transactions of the Royal Society of Edinburgh: Earth Sciences. 90 (4): 351–375. doi:10.1017/S0263593300002674.
- Shear, William A.; Edgecombe, Gregory D. (2010). "The geological record and phylogeny of the Myriapoda". Arthropod Structure & Development. 39 (2–3): 174–190. doi:10.1016/j.asd.2009.11.002. PMID 19944188.
- Edgecombe GD; Giribet G (2002). "Myriapod phylogeny and the relationships of Chilopoda" (PDF). In J Llorente Bousquets; JJ Morrone; HP Ulloa (eds.). Biodiversidad, Taxonomia y Biogeografia de Artropodos de Mexico: Hacia una S´ıntesis de su Conocimiento. Univ. Nac. Aut´on Mexico: Prensas Ciencias. pp. 143–168.
- Miyazawa, Hideyuki; Ueda, Chiaki; Yahata, Kensuke; Su, Zhi-Hui (2014). "Molecular phylogeny of Myriapoda provides insights into evolutionary patterns of the mode in post-embryonic development". Scientific Reports. 4 (4127): 4127. Bibcode:2014NatSR...4E4127M. doi:10.1038/srep04127. PMC 3927213. PMID 24535281.
- Zwick, Andreas; Regier, Jerome C.; Zwickl, Derrick J.; Gadagkar, Sudhindra R. (2012). "Resolving Discrepancy between Nucleotides and Amino Acids in Deep-Level Arthropod Phylogenomics: Differentiating Serine Codons in 21-Amino-Acid Models". PLOS ONE. 7 (11): e47450. Bibcode:2012PLoSO...747450Z. doi:10.1371/journal.pone.0047450. PMC 3502419. PMID 23185239.
External links
| Wikispecies has information related to Myriapoda. |
| Wikimedia Commons has media related to Myriapoda. |