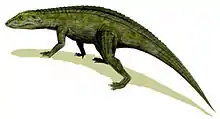
Ornithosuchidae
Ornithosuchidae is an extinct family of pseudosuchian archosaurs (distant relatives of modern crocodilians) from the Triassic period. Ornithosuchids were quadrupedal and facultatively bipedal (i.e. chimpanzees), meaning that they had the ability to walk on two legs for short periods of time. They had distinctive, downturned snouts, unique, "crocodile-reversed" ankle bones, and several other features that distinguish them from other archosaurs. Ornithosuchids were geographically widespread during the Carnian and Norian stages of the Late Triassic with members known from Argentina, Brazil, and the United Kingdom. Four genera, comprising Ornithosuchus, Venaticosuchus, Dynamosuchus,[1] and Riojasuchus are presently known.[2] The family was first erected by German paleontologist Friedrich von Huene in 1908.[3]
| Ornithosuchidae | |
|---|---|
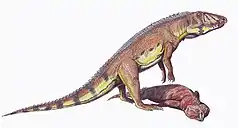 | |
| Life restoration of Ornithosuchus scavenging on the rhynchosaur Hyperodapedon | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Pseudosuchia |
| Family: | †Ornithosuchidae von Huene, 1908 |
| Type species | |
| †Ornithosuchus woodwardi Newton, 1894 | |
| Genera | |
|
†Aenigmaspina? | |
Description
Skull
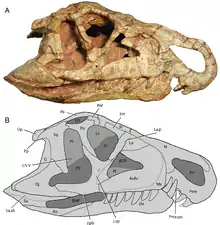
Ornithosuchids can be identified by the presence of an arched diastema, a gap between the teeth at the front of the snout. When the jaw is closed, two large curved dentary (lower jaw) teeth fit into the diastema, which is positioned between the premaxilla and maxilla. There are two shallow depressions on the wall of the diastema to accommodate these teeth. The large dentary teeth of Ornithosuchus and Riojasuchus are placed behind a smaller procumbent dentary tooth that sticks out from the jaw. This type of tooth position is not seen in any other basal archosaurs. Another characteristic feature of ornithosuchids is their unusual downturned, overhanging snout, seen in Riojasuchus and Venaticosuchus, but not Ornithosuchus.[4]
Several other features distinguish ornithosuchids from all other early archosaurs. Ornithosuchus and Riojasuchus both possess a small fenestra, or hole, between the palatine and pterygoid bones of the palate, i.e. the roof of the mouth. The contact between the nasal and prefrontal bones of the skull is small or absent, excluded by a large contact between the frontals and lacrimals. In other archosaurs, including rauisuchians, aetosaurs, pterosaurs, and dinosauromorphs, the nasal-prefrontal contact separates the frontal from the lacrimal.[4]
Postcranial skeleton
The postcranial skeleton is nearly completely known in Riojasuchus, incomplete in Ornithosuchus, and entirely unknown in Venaticosuchus. As a result, it is uncertain whether all of the postcranial traits seemingly unique to ornithosuchids actually occurred in all members of the family. Ornithosuchids known from decent postcranial remains typically had about 9 cervical (neck), 14-15 dorsal (back), 3 sacral (hip), and over 20 caudal (tail) vertebrae. Above each vertebra was a pair of bony scutes known as osteoderms.[2]
The femur (thigh bone) has a pronounced anterior trochanter. The anterior trochanter, sometimes known as the "lesser trochanter" (but unrelated to the lesser trochanter of the femur in humans), is a ridge on the outer surface of the femur, near the femoral head. It was probably an insertion point for the iliofemoralis cranialis muscle which helps to raise the leg. Most archosaurs and archosaur relatives lack a distinct anterior trochanter, but ornithosuchids are an exception, along with most dinosauromorphs (dinosaurs and their close relatives).[5]
Much like the femur, the fibula (outer shin bone) also has a distinctive point for muscle insertion. The muscle in question is the iliofibularis, which helps to straighten the limbs. In most archosaurs, the iliofibularis is inserted onto the fibula by means of a tiny ridge on the proximal part of the fibula, near the knee. However, ornithosuchids have a much larger, knob-shaped iliofibularis insertion point located about midway down the shaft of the fibula. Phytosaurs and aetosaurs also share a knob-like attachment point midway down the fibula, so it is unclear whether the case in ornithosuchids is a unique case of convergent evolution, or alternatively the retention of a trait independently lost by several archosaur lineages.[6]
Unlike most other early archosaurs, the pedal unguals (the distalmost bones of the feet that form claws) are laterally compressed. They are sharp and recurved. The unguals are very deep, being taller than they are long, especially on the inner digits. This type of claw is not seen in any other Triassic archosaur except for pterosaurs.[4]

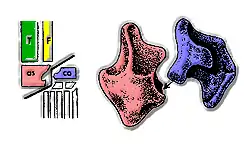
Major archosaur groups have often been distinguished from each other based on the structure of their ankles. In most crurotarsans, the astragalus has a convex projection that fits into a concave space in the calcaneum. This condition is often referred to as a "crocodile-normal" ankle, as it is the most common ankle type in crurotarsans. Ornithosuchids are unique among crurotarsans, and all other archosaurs, in their possession of a "crocodile-reversed" ankle. In a "crocodile-reversed" ankle, the placement of the concavity is reversed: instead of being on the calcaneum, it is on the astragalus. In ornithosuchids, the calcaneum bears a convex projection that is analogous to the convex projection on the "crocodile-normal" astragalus.[4]
Phylogeny
Ornithosuchidae is generally considered to be within the larger clade Suchia, which includes aetosaurs, rauisuchians, and crocodylomorphs.[7][8][9] Below is a cladogram based on Nesbitt (2011), showing the placement of Ornithosuchidae in Archosauriformes.[6]
| Archosauriformes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
References
- Müller, Rodrigo T.; Von Baczko, M. Belén; Desojo, Julia B.; Nesbitt, Sterling J. (31 January 2020). "The first ornithosuchid from Brazil and its macroevolutionary and phylogenetic implications for Late Triassic faunas in Gondwana" (PDF). Acta Palaeontologica Polonica. 65. doi:10.4202/app.00652.2019.
- Baczko, M. Belén von; Ezcurra, Martín D. (2013-01-01). "Ornithosuchidae: a group of Triassic archosaurs with a unique ankle joint". Geological Society, London, Special Publications. 379 (1): 187–202. doi:10.1144/SP379.4. ISSN 0305-8719.
- Huene, F. von. 1908. "Die Dinosaurier der europäischen Triasformation mit Berücksichtigung der aussereuropäischen Vorkommnisse". Geologische und Paläontologische Abhandlungen 1(Suppl.): 1–419
- Sereno, P.C. (1991). "Basal archosaurs: phylogenetic relationships and functional implications". Journal of Vertebrate Paleontology. 11 (Suppl. 4): 1–53. doi:10.1080/02724634.1991.10011426.
- Langer, Max C.; Benton, Michael J. (6 November 2006). "Early dinosaurs: A phylogenetic study" (PDF). Journal of Systematic Palaeontology. 4 (4): 309–358. doi:10.1017/s1477201906001970. ISSN 1477-2019.
- Nesbitt, S.J. (2011). "The early evolution of archosaurs: relationships and the origin of major clades" (PDF). Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1. hdl:2246/6112.
- Nesbitt, S.J.; Norrell, M.A. (2006). "Extreme convergence in the body plans of an early suchian (Archosauria) and ornithomimid dinosaurs (Theropoda)". Proceedings of the Royal Society B. 273 (1590): 1045–1048. doi:10.1098/rspb.2005.3426. PMC 1560254. PMID 16600879.
- Nesbitt, S.J. (2007). "The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa" (PDF). Bulletin of the American Museum of Natural History. 302: 1–84. doi:10.1206/0003-0090(2007)302[1:TAOEOA]2.0.CO;2. hdl:2246/5840.
- Brusatte, S.L.; Benton, M.J.; Desojo, J.B.; Langer, M.C. (2010). "The higher-level phylogeny of Archosauria (Tetrapoda: Diapsida)" (PDF). Journal of Systematic Palaeontology. 8 (1): 3–47. doi:10.1080/14772010903537732.
External links
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||