Rauisuchia
"Rauisuchia" is a group of mostly large (often 4 to 6 metres (13 to 20 ft)) Triassic archosaurs. It belongs to a larger clade called Pseudosuchia, which encompasses all archosaurs more closely related to crocodilians than to birds and other dinosaurs. "Rauisuchia" is currently considered an evolutionary grade, or even a wastebin taxon. It includes most of the large, carnivorous pseudosuchians that lived during the Triassic Period. Since crocodylomorphs likely originated from an ancestor that would have been a "rauisuchian", Rauisuchia in its traditional sense is considered paraphyletic as it excludes crocodylomorphs. To designate it as an informal group in scientific literature, the name in its traditional sense is often enclosed in quotation marks.[2] The monophyletic equivalent of the group Rauisuchia is the clade Paracrocodylomorpha. Paracrocodylomorpha consists of two branches: Poposauroidea, which includes a variety of strange archosaurs, and Loricata, which includes traditional "rauisuchians" and their crocodylomorph descendants.[2]
| "Rauisuchians" | |
|---|---|
 | |
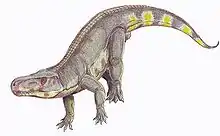
| Mounted skeleton of Prestosuchus chiniquensis (a basal loricatan) in the American Museum of Natural History. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Suchia |
| Informal group: | †Rauisuchia Huene, 1942 |
| Subgroups | |
Characteristics
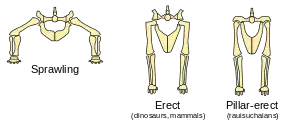

"Rauisuchians" had an erect gait with their legs oriented vertically beneath the body rather than sprawling outward. This type of gait is also seen in dinosaurs, but evolved independently in the two groups. In dinosaurs, the hip socket faces outward and the femur (thigh bone) connects to the side of the hip; while in rauisuchians, the hip socket faces downward to form a shelf of bone under which the femur connects.[3][4] This has been referred to as the pillar-erect posture.[5]
"Rauisuchians" lived throughout most of the Triassic. Along with many other large archosaurs, the group died out in the Triassic-Jurassic extinction event (barring crocodylomorphs, which survive to the present in the form of crocodilians). After their extinction, theropod dinosaurs were able to emerge as the sole large terrestrial predators, though there is still some debate over how the extinction influenced dinosaur evolution. The footprints of meat-eating dinosaurs may have suddenly increased in size at the start of the Jurassic, when rauisuchians were absent.[6] However, the apparent increase in dinosaur footprint size has instead been argued to be a result of increasing abundance of large theropods, rather than an abrupt acquisition of large size.[7] Some "rauisuchians" may have existed in the very early Jurassic based on bone fragments from South Africa, but this identification is tentative.[1]
The name "Rauisuchia" comes from the genus Rauisuchus, which was named after fossil collector Dr. Wilhelm Rau. The name Rauisuchus means Wilhelm Rau's crocodile.
Fossil record
Well-known "rauisuchians" include Ticinosuchus of the Middle Triassic of Europe (Switzerland and Northern Italy), Saurosuchus of the late Triassic (Late Carnian) of South America (Argentina), and Postosuchus of the late Triassic (Late Carnian to Early Norian) of North America (SW USA). One "rauisuchian", Teratosaurus, was for a long time even considered an early theropod dinosaur,[8] but was later shown to be non-dinosaurian.[9][10]
History of classification
"Rauisuchians" were originally thought to be related to erythrosuchids,[11] but it is now known that they are pseudosuchians.[12][2] Three families have historically been recognised: Prestosuchidae, Rauisuchidae, and Poposauridae, as well as a number of forms (e.g. those from the Olenekian of Russia) that are too primitive and/or poorly known to fit in any of these groups.
There has been considerable suggestion that the group as currently defined is paraphyletic, representing a number of related lineages independently evolving and filling the same ecological niche of medium to top terrestrial predator. For example, Parrish (1993)[13] and Juul (1994)[14] considered poposaurid rauisuchians to be more closely related to Crocodilia than to prestosuchids. Nesbitt (2003)[15] presented a different phylogeny with a monophyletic Rauisuchia. The group may even be something of a "wastebasket taxon". Determining exact phylogenetic relationships is difficult because of the scrappy nature of a lot of the material. However, further discoveries and studies, such as a study on the braincase of Batrachotomus (2002)[16] and restudies of other forms, such as Erpetosuchus (2002)[17] have shedding light on the evolutionary relationships of this poorly known group.
Cladistics
Despite its inclusion as an informal grouping in numerous phylogenetic studies, "Rauisuchia" has never received a formal definition. Most analyses in the past decade have found "Rauisuchia" to be a paraphyletic grouping, including all studies with a large sample size. Those that found the possibility that it was a natural group produced only weak support for this hypothesis.[18] In his large 2011 analysis of archosaurian relationships, Nesbitt recommended that the term "Rauisuchia" be abandoned.[2]
In a study of the ctenosauriscid Arizonasaurus, paleontologist Sterling Nesbitt defined a clade of rauisuchians called "Group X".[19] This group includes Arizonasuchus, Lotosaurus, Sillosuchus, Shuvosaurus, and Effigia. One distinguishing feature of Group X is their lack of osteoderms, which are common among many other crurotarsans. Many more features are found in the pelvis, including fully fused sacral vertebrae and a long, thin crest on the illium called the supra-acetabular crest. Additionally, many members of Group X have smooth frontal and nasal bones, which make up the upper portion of the rostrum. In other "rauisuchians" and many other crurotarsans, this area has bumps and ridges.[20] "Group X" is now termed Poposauroidea.[2]
Nesbitt later erected another clade, "Group Y", in 2007.[20] Group Y falls within Group X to include Sillosuchus, Shuvosaurus, and Effigia. Group Y is diagnosed by the presence of four or more sacral vertebrae with fully fused neural arches, which is also seen in theropod dinosaurs (a case of evolutionary convergence). In addition, the cervical vertebrae that make up the neck are strongly amphicoelus, meaning that they are concave at both ends. The fourth trochanter, a ridge of bone on the femur for muscle attachment seen in nearly all archosaurs, is absent in Group Y.[20] "Group Y" is now termed Shuvosauridae.[2]
Although not placed within Group Y, Lotosaurus shares many similarities with members of the clade, foremost of which is edentulous, or toothless, jaws. Edentulism is also seen in Shuvosaurus and Effigia, which have beak-like jaws. Nesbitt suggested that the derived characters of Lotosaurus may indicate that it is a transitional form between basal members of Group X and members of Group Y.[20]
Below is the cladogram from Nesbitt (2007):[20]
| Rauisuchia |
| ||||||||||||||||||||||||||||||||||||
In their phylogenetic study of archosaurs, Brusatte et al. (2010) found only weak support for Rauisuchia as a monophyletic grouping. As a result of their analysis, two clades were found to be within Rauisuchia, which they named Rauisuchoidea and Poposauroidea. Rauisuchoidea included Rauisuchidae and Prestosuchidae, as well as several basal taxa that were once assigned to the families, including Fasolasuchus and Ticinosuchus. Poposauroidea included poposaurids and ctenosauriscids, but the phylogeny had a large polytomy of genera in both groups that was difficult to resolve, which included Arizonasaurus, Poposaurus, and Sillosuchus. However, the characters linking these two groups were weak, and the question as to whether or not "Rauisuchia" forms a natural group remains unresolved.[18] Brusatte et al. (2010) was one of the last studies to find a monophyletic Rauisuchia clade.
Below is the cladogram from Brusatte et al. (2010):[18]
| Rauisuchia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In a more thorough test of archosaurian relationships published in 2011 by Sterling Nesbitt, "rauisuchians" were found to be paraphyletic, with Poposauroidea at the base of the clade Paracrocodylomorpha, and the rest of the "rauisuchians" forming a grade within the clade Loricata. Nesbitt noted that no previous study of "rauisuchian" relationships had ever included a wide variety of supposed "rauisuchians" as well as a large number of non-"rauisuchian" taxa as controls.[2]
Notes
- Tolchard, F.; Nesbitt, S. J.; Desojo, J. B.; Viglietti, P.; Butler, R. J.; Choiniere, J. N. (2019). "'Rauisuchian' material from the lower Elliot Formation of South Africa and Lesotho: Implications for Late Triassic biogeography and biostratigraphy". Journal of African Earth Sciences. 160 (103610): 103610. Bibcode:2019JAfES.16003610T. doi:10.1016/j.jafrearsci.2019.103610. Retrieved 10 November 2019.
- Nesbitt, S.J. (2011). "The early evolution of archosaurs: relationships and the origin of major clades" (PDF). Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1. hdl:2246/6112. S2CID 83493714.
- Bonaparte, J. F. (1984). "Locomotion in rauisuchid thecodonts". Journal of Vertebrate Paleontology. 3 (4): 210–218. doi:10.1080/02724634.1984.10011976.
- Benton, M. J. (1984). "Rauisuchians and the success of dinosaurs". Nature. 310 (5973): 101. doi:10.1038/310101a0. S2CID 4322293.
- Benton, M. J.; Clark, J. (1988). "Archosaur phylogeny and the relationships of the Crocodylia". In Benton, M.J. (ed.). The Phylogeny and Classification of the Tetrapods. 1. Amphibians, Reptiles, Birds. Systematics Association. 35A. Oxford: Clarendon Press. pp. 289–332.
- Olsen, P. E.; Kent, D. V.; Sues, H.-D.; Koeberl, C.; Huber, H.; Montanari, E. C.; Rainforth, A.; Fowell; S. J.; et al. (2002). "Ascent of Dinosaurs Linked to an Iridium Anomaly at the Triassic-Jurassic Boundary". Science. 296 (5571): 1305–1307. Bibcode:2002Sci...296.1305O. doi:10.1126/science.1065522. PMID 12016313. S2CID 24911506.
- Griffin, Christopher T. (September 2019). "Large neotheropods from the Upper Triassic of North America and the early evolution of large theropod body sizes". Journal of Paleontology. 93 (5): 1010–1030. doi:10.1017/jpa.2019.13. ISSN 0022-3360.
- See for example Colbert, E.H., 1961, Dinosaurs: Their Discovery and Their World, Dutton, New York, 1961 p.67
- Galton, P. M. (1985). "The poposaurid thecodontian Teratosaurus suevicus von Meyer, plus referred specimens mostly based on prosauropod dinosaurs". Stuttgarter Beiträge zur Naturkunde B. 116: 1–29.
- Benton, M.J. (1986). "The late Triassic reptile Teratosaurus - a rauisuchian, not a dinosaur". Palaeontology. 29: 293–301.
- Sill, W. D. (1974). "The anatomy of Saurosuchus galilei and the relationships of the rauisuchid thecodonts". Bulletin of the Museum of Comparative Zoology. 146: 317–362. ISSN 0027-4100.
- Benton, M. J. (2004). Vertebrate Paleontology (3rd ed.). Oxford: Blackwell Science. ISBN 0-632-05637-1.
- Parrish, J. M. (1993). "Phylogeny of the Crocodylotarsi, with reference to archosaurian and crurotarsan monophyly". Journal of Vertebrate Paleontology. 13 (3): 287–308. doi:10.1080/02724634.1993.10011511. S2CID 84288744.
- Juul, L. (1994). "The phylogeny of basal archosaurs". Palaeontologia Africana. 31: 1–38.
- Nesbitt, S. J. (2003). "Arizonasaurus and its implications for archosaur divergence". Proceedings of the Royal Society B: Biological Sciences. 270 (Suppl. 2): S234–S237. doi:10.1098/rsbl.2003.0066. ISSN 0962-8452. PMC 1809943. PMID 14667392.
- Gower, D. J. (2002). "Braincase evolution in suchian archosaurs (Reptilia: Diapsida): evidence from the rauisuchian Batrachotomus kupferzellensis". Zoological Journal of the Linnean Society. 136 (1): 49–76. doi:10.1046/j.1096-3642.2002.00025.x.
- Benton, M. J.; Walker, A. D. (2002). "Erpetosuchus, a crocodile-like basal archosaur from the Late Triassic of Elgin, Scotland". Zoological Journal of the Linnean Society. 136 (1): 25–47. doi:10.1046/j.1096-3642.2002.00024.x. ISSN 0024-4082.
- Brusatte, S. L.; Benton, M. J.; Desojo, J. B.; Langer, M. C. (2010). "The higher-level phylogeny of Archosauria (Tetrapoda: Diapsida)" (PDF). Journal of Systematic Palaeontology. 8 (1): 3–47. doi:10.1080/14772010903537732. S2CID 59148006.
- Nesbitt, S.J. (2005). "Osteology of the Middle Triassic pseudosuchian archosaur Arizonasaurus babbitti". Historical Biology. 17 (1): 19–47. doi:10.1080/08912960500476499. S2CID 84326151.
- Nesbitt, S.J. (2007). "The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa" (PDF). Bulletin of the American Museum of Natural History. 302: 1–84. doi:10.1206/0003-0090(2007)302[1:TAOEOA]2.0.CO;2. hdl:2246/5840. ISSN 0003-0090.
References
- Carroll, R. L. (1988). Vertebrate Paleontology and Evolution. New York: WH Freeman & Co. ISBN 0-7167-1822-7.
- "Archosauromorpha: Rauisuchiformes". Palaeos. Archived from the original on 2009-10-21. Retrieved 11 January 2010.
- Poling, Jeff; Creisler, Ben (16 December 1996). "Rauisuchia Translation and Pronunciation Guide". Dinosauria On-Line. Archived from the original on 2009-07-13. Retrieved 11 January 2010.
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||