Centrosaurinae
Centrosaurinae (Greek: pointed lizards) is a subfamily of ceratopsid dinosaurs, a group of large quadrupedal ornithiscians. Centrosaurine fossil remains are known primarily from the northern region of Laramidia (modern day Alberta, Montana, and Alaska) but isolated taxa have been found in China and Utah as well.[1]
| Centrosaurines | |
|---|---|
 | |
| Centrosaurus "nasicornus" skeleton, Palaeontological Museum Munich | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | †Ornithischia |
| Family: | †Ceratopsidae |
| Subfamily: | †Centrosaurinae Lambe, 1915 |
| Type species | |
| †Centrosaurus apertus Lambe, 1904 | |
| Subgroups | |
| |
| Synonyms | |
| |
Defining features of centrosaurines include a large nasal horn, short supratemporal horns, and an ornamented frill projecting from the back of the skull.[2] With the exception of Centrosaurus apertus, all adult centrosaurines have spike-like ornaments midway up the skull.[3] Morphometric analysis shows that centrosaurines differ from other ceratopsian groups in skull, snout, and frill shapes.[4] There is evidence to suggest that male centrosaurines had an extended period of adolescence and sexual ornamention did not appear until adulthood.[2]
Centrosaurinae was named by paleontologist Lawrence Lambe in 1915, with Centrosaurus as the type genus. The centrosaurines are further divided into three tribes: the Nasutoceratopsini, the Centrosaurini, and the Pachyrhinosaurini by Ryan et al (2016).[5] Nasutoceratopsins are defined as centrosaurines closer to Nasutoceratops titusi than to Centrosaurus apertus and centrosaurins are defined as centrosaurines (more specifically eucentrosaurans) closer to Centrosaurus apertus than to Pachyrhinosaurus canadensis. The only division used up until then was Pachyrhinosaurini which was defined as centrosaurines closer to Pachyrhinosaurus canadensis than to Centrosaurus apertus.
Classification
The classification of centrosaurines and the relationships among the various species is complicated by a wide degree of variation between individuals and growth stages. Some features that have traditionally been used to classify these dinosaurs, like the number and arrangement of frill ornaments or spikes, have been discovered to be more variable than previously thought. For example, the cladogram presented below follows a 2016 phylogenetic analysis by Chiba et al. (2017).[6] These authors treated the species Rubeosaurus ovatus as distinct from Styracosaurus albertensis, and recovered several distinct clades within Centrosaurini, which together formed a sister group to the Pachyrhinosaurini:
| Centrosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
However, subsequent studies have cast doubt on the usefulness of minor variations in frill spike arrangement for classifying centrosaurines. In particular, large sample sizes of the species Centrosaurus apertus and Syracosaurus albertensis have shown a higher than predicted amount of variation. In 2020, Holmes et al. explored what the effect of recognizing such diversity would have on centrosaur classification. They used the same data as Chiba et al.'s 2017 study, but treated Rubeosaurus as a synonym of Styracosaurus, dropping it from their taxon list. The resulting cladogram (below) found Centrosaurini as a polytomy, a grouping with no discernable sister group relationships within it. The authors concluded that this meant the variation present within these species made it difficult to find any real resolution among them, and may even provide support for the hypothesis that centrosaurines evolved primarily via anagenesis (a single lineage changing through time) rather than cladogenesis (multiple branching lineages with shared common ancestors).[7]
| Centrosaurinae |
| ||||||||||||||||||||||||||||||||||||
}}
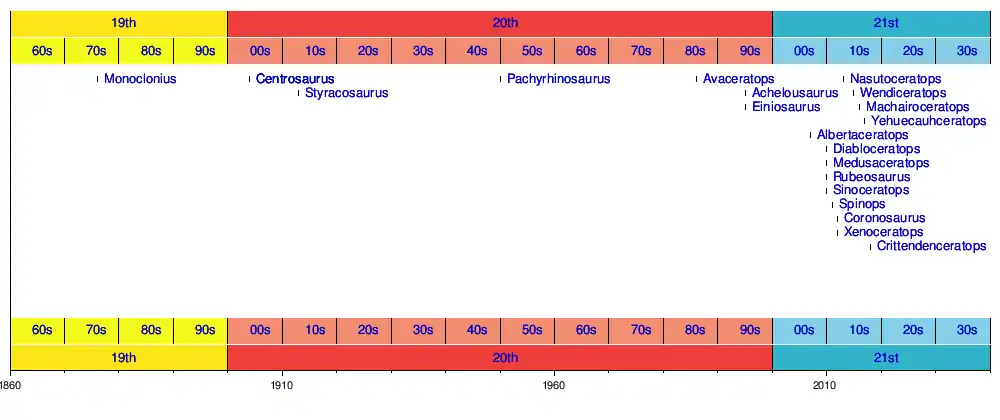
Biogeography

Centrosaurine fossils have mostly been found in Western North America (Alberta, Montana, & Alaska).[1] In the United States, two taxa, Diabloceratops and Machairoceratops, have been found as far south as Utah. Yehuecauhceratops is a nasutoceratopsin from Coahuila, Mexico and the southernmost occurrence of a centrosaurine in North America. [1] No centrosaurine fossils had been uncovered outside Western North America until 2010 when Sinoceratops zhuchengensis was discovered in the Shandong Province of China.[8] Some authors question the placement of Sinoceratops within Centrosaurinae, however. All other Late Cretaceous dinosaur groups from North America have been found in Asia as well so the initial absence of Asian centrosaurines had been surprising.[8] The current evidence suggests that Centrosaurinae originated in Laramidia 90-80 million years ago.[1] This means Sinoceratops would have migrated to China from North America.[5] Some hypothesize that centrosaurines originated in the south Laramidia and only later radiated north.[9]
Body size
Compared to their sister group, Chasmosaurinae, centrosaurines are relatively small. The primitive Sinoceratops zhuchengensis is an exception, with an estimated skull length of 180 cm.[8] By contrast, the skull length of Albertoceratops was more typical for this group at only 67 cm.[3] In general, centrosaurines were about the size of a rhinoceros with body lengths ranging from 2.5 to 8 meters.[10]
Reproduction
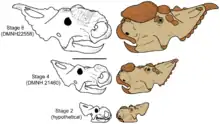
Possible neonate sized centrosaurine fossils have been documented in the scientific literature.[11] Research indicates that centrosaurines did not achieve fully developed mating signals until nearly fully grown.[12][2] Scott D. Sampson finds commonality between the slow growth of mating signals in centrosaurines and the extended adolescence of animals whose social structures are ranked hierarchies founded on age-related differences.[12] In these sorts of groups young males are typically sexually mature for several years before actually beginning to breed, when their mating signals are most fully developed.[13] Females, by contrast do not have such an extended adolescence.[13]
See also
Footnotes
- Sampson, Scott D.; Lund, Eric K.; Loewen, Mark A.; Farke, Andrew A.; Clayton, Katherine E. (2013-09-07). "A remarkable short-snouted horned dinosaur from the Late Cretaceous (late Campanian) of southern Laramidia". Proc. R. Soc. B. 280 (1766): 20131186. doi:10.1098/rspb.2013.1186. ISSN 0962-8452. PMC 3730592. PMID 23864598.
- Sampson, Scott D.; Ryan, Michael J.; Tanke, Darren H. (1997-11-01). "Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): taxonomic and behavioral implications". Zoological Journal of the Linnean Society. 121 (3): 293–337. doi:10.1111/j.1096-3642.1997.tb00340.x. ISSN 0024-4082.
- Ryan, Michael J. (2007-03-01). "A new basal centrosaurine ceratopsid from the oldman formation, southeastern alberta". Journal of Paleontology. 81 (2): 376–396. doi:10.1666/0022-3360(2007)81[376:ANBCCF]2.0.CO;2. ISSN 0022-3360.
- Maiorino, Leonardo; Farke, Andrew A; Kotsakis, Tassos; Piras, Paolo (2017). "Macroevolutionary patterns in cranial and lower jaw shape of ceratopsian dinosaurs (Dinosauria, Ornithischia): phylogeny, morphological integration, and evolutionary rates" (PDF). Evolutionary Ecology Research. 18: 123–167.
- Ryan, Michael J.; Holmes, Robert; Mallon, Jordan; Loewen, Mark; Evans, David C. (2016-10-27). "A basal ceratopsid (Centrosaurinae: Nasutoceratopsini) from the Oldman Formation (Campanian) of Alberta, Canada". Canadian Journal of Earth Sciences. 54 (1): 1–14. doi:10.1139/cjes-2016-0110. ISSN 0008-4077.
- Kentaro Chiba; Michael J. Ryan; Federico Fanti; Mark A. Loewen; David C. Evans (2018). "New material and systematic re-evaluation of Medusaceratops lokii (Dinosauria, Ceratopsidae) from the Judith River Formation (Campanian, Montana)". Journal of Paleontology. in press (2): 272–288. doi:10.1017/jpa.2017.62.
- Holmes, R. B., Persons, W. S., Rupal, B. S., Qureshi, A. J., & Currie, P. J. (2020). Morphological variation and asymmetrical development in the skull of Styracosaurus albertensis. Cretaceous Research, 107, 104308.
- Xu, Xing; Wang, KeBai; Zhao, XiJin; Li, DunJing (2010-06-01). "First ceratopsid dinosaur from China and its biogeographical implications". Chinese Science Bulletin. 55 (16): 1631–1635. doi:10.1007/s11434-009-3614-5. ISSN 1001-6538.
- Lund, Eric K.; O’Connor, Patrick M.; Loewen, Mark A.; Jinnah, Zubair A. (2016-05-18). "A New Centrosaurine Ceratopsid, Machairoceratops cronusi gen et sp. nov., from the Upper Sand Member of the Wahweap Formation (Middle Campanian), Southern Utah". PLOS ONE. 11 (5): e0154403. doi:10.1371/journal.pone.0154403. ISSN 1932-6203. PMC 4871575. PMID 27192148.
- V., Rey, Luis (2007). Dinosaurs : the most complete, up-to-date encyclopedia for dinosaur lovers of all ages. Random House. ISBN 9780375824197. OCLC 930042495.
- "Abstract," Tanke and Brett-Surman (2001). Page 207.
- "Retarded Growth of Mating Signals," Sampson (2001); page 270.
- "Sociological Correlates in Extant Vertebrates," Sampson (2001); page 265.
References
- Sampson, S. D. (1995b). "Two new horned dinosaurs from the Upper Cretaceous Two Medicine Formation of Montana; with a phylogenetic analysis of the Centrosaurinae (Ornithischia: Ceratopsidae)". Journal of Vertebrate Paleontology. 15 (4): 743–760. doi:10.1080/02724634.1995.10011259.
- Sampson, S. D., 2001, Speculations on the socioecology of Ceratopsid dinosaurs (Orinthischia: Neoceratopsia): In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 263–276.
- Tanke, D.H. and Brett-Surman, M.K. 2001. Evidence of Hatchling and Nestling-Size Hadrosaurs (Reptilia:Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta, Canada. pp. 206–218. In: Mesozoic Vertebrate Life—New Research Inspired by the Paleontology of Philip J. Currie. Edited by D.H. Tanke and K. Carpenter. Indiana University Press: Bloomington. xviii + 577 pp.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||





