Enoyl-CoA hydratase
Enoyl-CoA hydratase (ECH) or crotonase[1] is an enzyme that hydrates the double bond between the second and third carbons on 2-trans/cis-enoyl-CoA:[2]
| enoyl-Coenzyme A, hydratase/3-hydroxyacyl Coenzyme A dehydrogenase | |||||||
|---|---|---|---|---|---|---|---|
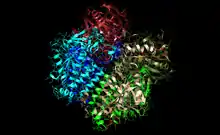 Enoyl-CoA hydratase hexamer from a rat with active site in orange and substrate in red. | |||||||
| Identifiers | |||||||
| Symbol | EHHADH | ||||||
| Alt. symbols | ECHD | ||||||
| NCBI gene | 1962 | ||||||
| HGNC | 3247 | ||||||
| OMIM | 607037 | ||||||
| RefSeq | NM_001966 | ||||||
| UniProt | Q08426 | ||||||
| Other data | |||||||
| EC number | 4.2.1.17 | ||||||
| Locus | Chr. 3 q26.3-q28 | ||||||
| |||||||
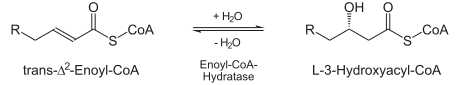
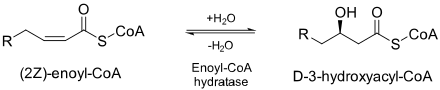
ECH is essential to metabolizing fatty acids in beta oxidation to produce both acetyl CoA and energy in the form of ATP.[2]
ECH of rats is a hexameric protein (this trait is not universal, but human enzyme is also hexameric), which leads to the efficiency of this enzyme as it has 6 active sites. This enzyme has been discovered to be highly efficient, and allows people to metabolize fatty acids into energy very quickly. In fact this enzyme is so efficient that the rate for short chain fatty acids is equivalent to that of diffusion-controlled reactions.[3]
Metabolism
Fatty acid metabolism
ECH catalyzes the second step (hydratation) in the breakdown of fatty acids (β-oxidation).[4] Fatty acid metabolism is how human bodies turn fats into energy. Fats in foods are generally in the form of triglycerols. These must be broken down in order for the fats to pass into human bodies. When that happens, three fatty acids are released.
Leucine metabolism
Mechanism
ECH is used in β-oxidation to add a hydroxyl group and a proton to the unsaturated β-carbon on a fatty-acyl CoA. ECH functions by providing two glutamate residues as catalytic acid and base. The two amino acids hold a water molecule in place, allowing it to attack in a syn addition to an α-β unsaturated acyl-CoA at the β-carbon. The α-carbon then grabs another proton, which completes the formation of the beta-hydroxy acyl-CoA.
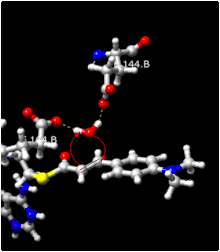
It is also known from experimental data that no other sources of protons reside in the active site. This means that the proton which the α-carbon grabs is from the water that just attacked the β-carbon. What this implies is that the hydroxyl group and the proton from water are both added from the same side of the double bond, a syn addition. This allows ECH to make an S stereoisomer from 2-trans-enoyl-CoA and an R stereoisomer from the 2-cis-enoyl-CoA. This is made possible by the two glutamate residues which hold the water in position directly adjacent to the α-β unsaturated double bond. This configuration requires that the active site for ECH is extremely rigid, to hold the water in a very specific configuration with regard to the acyl-CoA. The data for a mechanism for this reaction is not conclusive as to whether this reaction is concerted (shown in the picture) or occurs in consecutive steps. If occurring in consecutive steps, the intermediate is identical to that which would be generated from an E1cB-elimination reaction.[8]
ECH is mechanistically similar to fumarase.
References
- "EC 4.2.1.17". www.sbcs.qmul.ac.uk. Retrieved 2018-09-05.
- Allenbach, L; Poirier, Y (2000). "Analysis of the Alternative Pathways for the β-Oxidation of Unsaturated Fatty Acids Using Transgenic Plants Synthesizing Polyhydroxyalkanoates in Peroxisomes". Plant Physiology. 124 (3): 1159–1168. doi:10.1104/pp.124.3.1159. ISSN 0032-0889. PMC 59215. PMID 11080293.
- Engel CK, Kiema TR, Hiltunen JK, Wierenga RK (February 1998). "The crystal structure of enoyl-CoA hydratase complexed with octanoyl-CoA reveals the structural adaptations required for binding of a long chain fatty acid-CoA molecule". Journal of Molecular Biology. 275 (5): 847–59. doi:10.1006/jmbi.1997.1491. PMID 9480773.
- Cox DL, Nelson MM (2005). Lehninger principles of biochemistry (4th ed.). New York: W.H. Freeman. p. 647-43. ISBN 978-0-7167-4339-2.
- Wilson JM, Fitschen PJ, Campbell B, Wilson GJ, Zanchi N, Taylor L, Wilborn C, Kalman DS, Stout JR, Hoffman JR, Ziegenfuss TN, Lopez HL, Kreider RB, Smith-Ryan AE, Antonio J (February 2013). "International Society of Sports Nutrition Position Stand: beta-hydroxy-beta-methylbutyrate (HMB)". Journal of the International Society of Sports Nutrition. 10 (1): 6. doi:10.1186/1550-2783-10-6. PMC 3568064. PMID 23374455.
- Kohlmeier M (May 2015). "Leucine". Nutrient Metabolism: Structures, Functions, and Genes (2nd ed.). Academic Press. pp. 385–388. ISBN 978-0-12-387784-0. Retrieved 6 June 2016.
Energy fuel: Eventually, most Leu is broken down, providing about 6.0kcal/g. About 60% of ingested Leu is oxidized within a few hours ... Ketogenesis: A significant proportion (40% of an ingested dose) is converted into acetyl-CoA and thereby contributes to the synthesis of ketones, steroids, fatty acids, and other compounds
Figure 8.57: Metabolism of L-leucine - Bahnson BJ, Anderson VE, Petsko GA (February 2002). "Structural mechanism of enoyl-CoA hydratase: three atoms from a single water are added in either an E1cb stepwise or concerted fashion". Biochemistry. 41 (8): 2621–9. doi:10.1021/bi015844p. PMID 11851409.
External links
- Enoyl-CoA+Hydratase at the US National Library of Medicine Medical Subject Headings (MeSH)